
Статья опубликована в рамках: IV Международной научно-практической конференции «Научные достижения биологии, химии, физики» (Россия, г. Новосибирск, 01 февраля 2012 г.)
Наука: Химия
Секция: Биоорганическая химия
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов
МОЛЕКУЛЯРНАЯ И ФИЗИКО-ХИМИЧЕСКАЯ ХАРАКТЕРИСТИКА OMPF- И OMPC-ПОДОБНЫХ БЕЛКОВ НАРУЖНОЙ МЕМБРАНЫ YERSINIA RUCKERI
Чистюлин Дмитрий Константинович
аспирант 2-го года обучения, Тихоокеанский институт биоорганической химии ДВО РАН, г. Владивосток
e-mail:cdk27gmail.ru
Вакорина Татьяна Ивановна,
канд. хим. наук, науч. сотр., Тихоокеанский институт биоорганической химии ДВО РАН, г. Владивосток
Ким Наталья Юрьевна
науч. сотр., Тихоокеанский институт биоорганической химии ДВО РАН, г. Владивосток
Хоменко Валентина Александровна
канд. хим. наук, ст. науч. сотр., Тихоокеанский институт биоорганической химии ДВО РАН, г. Владивосток
Портнягина Ольга Юрьевна
канд. биол. наук, ст. науч. сотр., Тихоокеанский институт биоорганической химии ДВО РАН, г. Владивосток
Новикова Ольга Данииловна
д-р хим. наук, вед. науч. сотр., Тихоокеанский институт биоорганической химии ДВО РАН, г. Владивосток
Работа поддержана грантами по Программе Президиума РАН “Молекулярная и клеточная биология” и РФФИ – ДВО РАН № 11-04-98584.
Yersinia ruckeri – грамотрицательная бактерия, вызывающая иерсиниоз у рыб. Это септическое заболевание, поражающее лососевых, особенно - радужную форель, которое регистрируется в Прибалтийском регионе, в большинстве стран Восточной и Западной Европы и в США. К факторам, способствующим возникновению и распространению заболевания, относятся неблагоприятные условия окружающей среды (дефицит кислорода, накопление органических веществ, увеличение количества аммиака), а также высокие показатели плотности посадки рыб. Степень патогенности микроорганизмов, обитающих в водной среде, в значительной степени зависит от колебаний параметров последней, поскольку в данном случае внешние условия непосредственно влияют как на бактерии, так и на организм хозяина [7, 6]. Для бактерий, патогенных для рыб, регуляция температуры приобретает особое значение, поскольку регулирует экспрессию специфических белков, являющихся факторами патогенности. Экспрессия таких белков может уменьшаться (или прекращаться вовсе) при температуре, ниже оптимальной температуры роста бактерий [7, 6, 9]. В литературе существуют, например, данные о том, что синтез факторов вирулентности Y. ruckeri зависит от различных условий, в том числе температуры окружающей среды [8, 9].
Однако участие порообразующих белков наружной мембраны (НМ), являющихся эффекторами патогенеза, в развитии инфекции, вызываемой Y. ruckeri, пока не рассматривалось. Влияние условий культивирования Y. ruckeri на регуляцию экспрессии этих белков изучено также недостаточно. В наших предварительных исследованиях было обнаружено, что температура культивирования влияет на полипептидный профиль фракции порообразующих белков (поринов) НМ Y. ruckeri. В НМ “холодового” (4°С) варианта микроорганизма экспрессируются преимущественно OmpF-подобные порины, а в НМ “теплового” (37°С) варианта – OmpC-подобные белки [5].
Настоящая работа посвящена выделению и сравнительной физико-химической характеристике OmpF- и OmpC-подобных белков НМ Y. ruckeri.
Как известно [10], порины относятся к β-структурированным мембранным белкам, которые в НМ грамотрицательных бактерий существуют в виде тримеров, устойчивых к действию денатурирующих агентов и протеолитических ферментов. Порины связаны очень прочной (но нековалентной связью) с нерастворимым слоем пептидогликана (ПГ), разделяющего наружную и цитоплазматическую мембраны. Солюбилизация поринов может быть осуществлена только с помощью детергентов, при этом пространственная организация белков зависит от природы используемого солюбилизирующего агента и температуры, при которой проводится диссоциация комплекса ПГ- порин [4]. В связи с этим, для получения поринов в конформации, максимально приближенной к таковой в нативной мембране, большое значение имеет подбор условий извлечения этих белков из НМ бактерий.
Комплекс ПГ- порообразующие белки Y. ruckeri получали по методу Розенбуша [12]. Для удаления белков цитоплазматической мембраны микробные клетки предварительно экстрагировали саркозилатом натрия. Для очистки комплекса ПГ-пориновые белки от нуклеиновых кислот использовали мультиферментный комплекс из гепатопанкреаса промысловых видов крабов [1]. Для получения изолированных OmpF- и OmpC- поринов в тримерной форме комплекс ПГ-пориновые белки экстрагировали 1 % SDS и 0.5 М NаCl, и полученную фракцию очищали с помощью гель-хроматографии на Sephacryl S-300 в присутствии SDS. Согласно данным электрофореза в полиакриламидном геле в присутствии SDS (SDS, ПААГ-электрофореза) [11] кажущиеся мол. массы мономеров OmpF- и OmpC поринов Y. ruckeri, составили 37000 ± 1000 и 39000± 1000 Да соответственно.
Данные, полученные в ходе эксперимента по термоденатурации выделенных белков, свидетельствовали об их различной термоустойчивости (рис. 1). Заметная полоса в области мономера OmpF белка появлялась при температуре 70—80°С, полное превращение олигомеров в денатурированный мономер наблюдалось только при температуре 90°С. В случае OmpC порина диссоциация тримеров белка начиналась при 65°С и заканчивалась при 75°С. Таким образом, подобно неспецифическим поринам НМ других грамотрицательных бактерий, OmpF порин Y. ruckeri более устойчив к действию температуры, нежели порин OmpC типа. Ранее мы показали, что температурный интервал перехода тримеров в мономеры для поринов из ряда патогенных и непатогенных видов иерсиний составляет 50—70°С [2].Однако, критическая температура необратимого конформационного перехода порина OmpF Y. ruckeri оказалась на 20 градусов выше по сравнению с поринами других видов иерсиний. Этот факт представляет особый интерес, поскольку до сих пор в литературе не встречалось сообщений о столь высокой степени термоустойчивости среди поринов НМ энтеробактерий.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
|
|
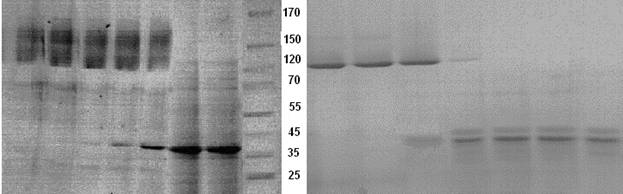
А Б
Рисунок 1. Термоденатурация поринов НМ Y. ruckeri. Инкубацию раствора белка проводили в интервале температур 60—100°С.
(А) исходный порин OmpF (1); порин инкубированный в течение 10 мин при 60°С (2), 70°С (3), 80°С (4), 85°С (5), 90°С (6), и100°С (7). Белки-маркеры (8).
(Б) исходный порин OmpC (9); порин инкубированный в течение 10 мин при 60°С (10), 65°С (11), 70°С (12), 75°С (13), 80°С (14), и100°С (15).
Нуклеотидные последовательности кодируемой части ompF и ompC генов Y. ruckeri (штамм КММ 821) были получены с помощью ПЦР амплификации с использованием ген-специфичных праймеров [5]. На основе этих последовательностей были выведены первичные структуры OmpF и OmpC-подобных поринов, топологические модели которых [14] приведены на рис. 2. Величины молекулярных масс мономеров OmpF – и OmpC- подобных поринов Y. ruckeri, рассчитанные из данных аминокислотной последовательности, составили 38015.20 и 39359.35 Да соответственно. Эти значения хорошо согласуются с величинами молекулярных масс, приведенными выше.
|
|
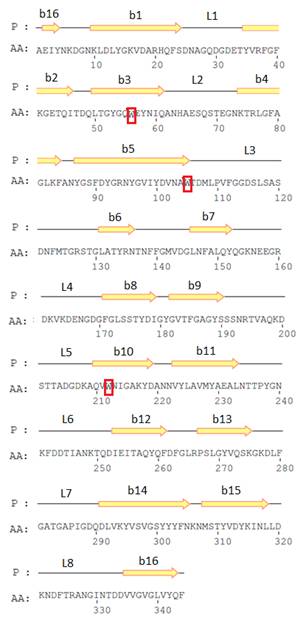
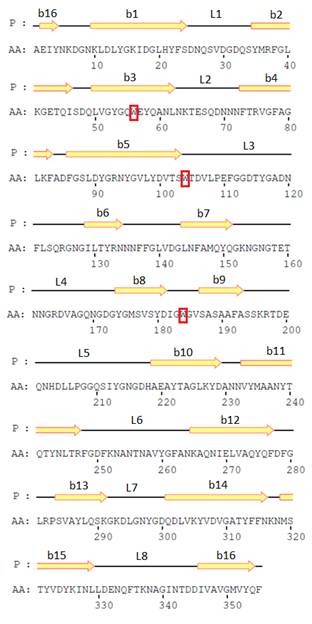
А Б
Рисунок 2. Топологические модели OmpF- и OmpC – поринов Y. ruckeri.
Стрелками обозначены участки аминокислотной последовательности, соответствующие β-стрэндам, линиями – участки наружных петель, внешних (L1—L8) и периплазматических. Отмечено также расположение остатков триптофана в полипептидной цепи белков.
Сравнительный анализ аминокислотных последовательностей исследуемых поринов и неспецифических поринов НМ Y. Pseudotuberculosis [3] показал, что степень гомологии между поринами OmpF- (или OmpC) типов разных видов иерсиний выше, чем между поринами разных типов одного вида микроорганизма. Так, степень гомологии аминокислотных последовательностей OmpF белков Y. ruckeri и Y. pseudotuberculosis составляет 61 %, OmpC белков этих видов иерсиний – 55 %, а степень гомологии OmpF и OmpC Y. ruckeri - только 48 %.
Для характеристики пространственной организации исследуемых белков в растворе использовали методы оптической спектроскопии: круговой дихроизм и собственную белковую флуоресценцию. Для солюбилизации образцов использовали 0.25 % раствор SDS, в котором, как было показано ранее [4], не происходит денатурации OmpF порина из псевдотуберкулезного микроба.
Круговой дихроизм. Спектр КД OmpF порина в ближней УФ-области (240—310 нм), области поглощения остатков ароматических хромофоров и дисульфидных связей, имеет достаточно выраженную тонкую структуру с положительной полосой при 275 нм, относящейся к остаткам тирозина, и «плечами» при 268 и 260 нм, относящимися к остаткам фенилаланина (рис. 3, А кривая 1).
|
|
|
|


Рисунок 3. Спектры КД OmpF (А) и OmpC (Б) НМ Y. ruckeri в ближней (250—300 нм) УФ-области (кривые 1) и дальней (190 —240 нм) УФ-области (кривые 2).
В спектре КД OmpC порина в этой УФ-области те же характеристичные полосы менее выражены и имеют меньшую эллиптичность (рис. 3, Б кривая 1). Полученные данные указывают на то, что третичная структура OmpF Y. ruckeri более жестко фиксирована в этих условиях, нежели структура OmpC белка.
Спектры КД исследуемых поринов в далекой УФ-области (190—240 нм), области поглощения пептидных связей, существенно различаются. OmpF белок имеет в этой области отрицательную полосу небольшой эллиптичности с «плечами» при 215 и 202 нм и положительную полосу малой эллиптичности при 195 нм (рис. 3, А кривая 2). Форма спектра КД этого порина указывает на достаточно высокое содержание β-структуры и присутствие небольшого количества α-спиральных участков в полипептидной цепи белка. Спектр КД OmpC порина в этой области имеет отрицательную полосу при 220 нм и положительную полосу большой эллиптичности при 197 нм, что свидетельствует о высоком содержании α-спирали во вторичной структуре белка. (рис. 3, Б кривая 2). Выводы, сделанные на основе качественной оценки спектров КД исследуемых поринов, были подтверждены расчётом элементов вторичной структуры (табл.), выполненным с помощью пакета программ CD Pro [13]. Как следует из данных таблицы, содержание α-спирали в OmpC порине в 3 раза больше, нежели в OmpF белке. Поскольку порообразующие белки обоих типов (OmpF и OmpC), выделенные из НМ Y. ruckeri имеют высокое содержание (62 и 60 %) суммарной β-структуры, их можно отнести к белкам β-класса, что характерно для изолированных поринов грамотрицательных бактерий в растворах детергентов.
Таблица. Содержание элементов вторичной структуры поринов OmpF и OmpC НМ Y. ruckeri, %
|
|
|
изгиб |
форма |
||||
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I, II, III –регулярная, искаженная и общая структуры соответственно.
Собственная белковая флуоресценция. Спектры суммарной и триптофановой флуоресценции исследуемых белков, имеют длинноволновое положение максимумов. Так, в случае Omp F порина максимум суммарной эмиссии находится при 335±1 нм, а максимум триптофановой флуоресценции – при 341±1 нм. Максимумы суммарной и триптофановой флуоресценции OmpC порина находятся, соответственно, при 339 и 340 нм. Этот факт, а также невысокая интенсивность флуоресценции остатков триптофана свидетельствуют о том, что значительная часть остатков этого ароматического флуорофора в составе обоих белков доступна растворителю. Это подтверждается аппроксимацией суммарной эмиссии белков спектральными формами излучения остатков триптофана и тирозина (рис. 4). Как видно из данных рис. 4, на долю спектральной формы III в случае OmpC порина приходится 45 %, в случае OmpF белка - более половины всех остатков триптофана.
|
|
|
|


Рисунок 4. Спектры флуоресценции поринов OmpF (А) и OmpC (Б)НМ Y. ruckeri и их аппроксимация спектральными формами излучения остатков тирозина (Y) и триптофана (формы S, I, III). Возбуждение 280 нм. Температура 20 оС.
А – Спектр OmpF порина: вклады остатков тирозина (9 %) и спектральных форм триптофана S, I и III (27.8, 5.1 и 58.1 % соответственно).
Б - Спектр OmpС порина: вклады остатков тирозина (11.4 %) и спектральных форм триптофана S, I и III (10.9, 32.4 и 45.3 % соответственно).
Таким образом, из НМ Y. ruckeri, выделены и охарактеризованы β-структурированные мембранные белки, которые по молекулярной массе и пространственной организации подобны неспецифическим поринам иерсиний [12, 2] и других энтеробактерий [4]. По сравнению с известными поринами энтеробактерий неспецифический OmpF белок НМ Y. ruckeri обладает повышенной устойчивостью по отношению к температуре.
Список литературы:
1. Артюков А. А., Мензорова Н. И., Козловская Э. П., Кофанова Н. Н., Козловский А. С., Рассказов В. А. Патент 2280076 РФ, МПК С12N 9/48, 9/64 (2006.01) Ферментативный препарат из гепатопанкреаса промысловых видов крабов и способ его получения; заявители и патентообладатели ТИБОХ ДВО РАН. — № 2004135771/13; Заявл.06.12.2004; Опубл. 20.07.2006, Бюл. № 20. 6 с.
2. Вострикова О. П., Ким Н. Ю., Лихацкая Г. Н., Гузев К. В., Вакорина Т. И., Хоменко В. А., Новикова О. Д., Соловьева Т. Ф. Структура и функция порообразующих белков бактерий рода Yersinia I. Выделение и сравнительная характеристика физико-химических свойств и функциональной активности поринов иерсиний // Биоорган. химия. 2006. Т. 32, № 4. С. 371—383.
3. Гузев К. В., Исаева М. П., Новикова О. Д., Соловьева Т. Ф., Рассказов В. А. Молекулярная характеристика OmpF-подобных поринов патогенных Yersinia // Биохимия. 2005. Т. 70, вып. 10. С. 1338—1345.
4. Новикова О. Д., Федореева Л. И., Хоменко В. А., Портнягина О. Ю., Ермак И. М., Лихацкая Г. Н., Мороз С. В., Соловьева Т. Ф., Оводов Ю. С. Влияние способа экстракции порообразующего белка из Yersinia pseudotuberculosis на его макромолекулярную организацию // Биоорган. химия. 1993. Т. 19, № 5. C. 536—547.
5. Чистюлин Д. К., Портнягина О. Ю., Хоменко В. А., Вакорина Т. И., Ким Н. Ю., Исаева М. П., Лихацкая Г. Н., Новикова О. Д.OmpF-подобный поринн наружной мембраны Yersinia ruckeri // В: Сб. тезисов докладов III Всероссийской научно—практической конференции с международным участием «Инфекции, обусловленные иерсиниями». Санкт-Петербург. 12—14 октября 2011 г. С. 114—115.
6. Altinok I. The infectious route of Yersinia ruckeri is affected by salinity. Bull. Eur. association Fish // Pathologists. 2004. V. 24. P. 253—259.
7. Altinok I., Grizzle J.M. Effects of salinity on Yersinia ruckeri infection of rainbow trout and brown trout // J. Aquatic Animal Health. 2011. V. 3. P. 334—339;
8. Fernandes L., Marques I., Guijarro J. A. Identification of specific in vivo-induced genes in Yersinia ruckeri and analysis of ruckerbactin, a catecholate siderophore iron acquisition system // Appl.Environ. Microbiol. 2004. V. 70. P. 5199—5207.
9. Fernandez L., Lopez J.R., Secades P., Menendez A., Marquez I. & Guijarro J. A. (In vitro and in vivo studies of the Yrp1 protease from Yersinia ruckeri and its role in protective immunity against enteric red mouth disease of salmonids. Appl. Environ. Microbiol. 2003. V. 69 P. 7328—7335.
10. Garavito R. M., Rosenbusch J. P. Isolation and crystallization of bacterial porin // Methods Enzymol. 1986. V. 125. P. 309—328.
11. Lugtenberg B., Meijers J., Peters R., van der Hoek P., van Alphen L. Electrophoretic resolution of the major outer membrane of Escherichia coli K-12 into four bands // FEBS Lett. 1975. V. 58. P. 254—258.
12. Rosenbusch J. P. Characterization of the major envelope protein from Escherichia coli. Regular arrangement on the peptidoglycan and unusual dodecyl sulfate binding // J. Biol. Chem. 1974.V. 249, P. 8019—8029.
13. Sreerama N., Woody R. W. Estimation protein secondary structure from circular dichroism spectra: comparison of CONTIN, SELCON and CDSSTR methods with an expanded reference set // Anal. Biochem. 2000. V. 287. P. 252—260.
14. The PSIPRED Protein Structure Prediction Server. [электронный ресурс] — Режим доступа. — URL: http://bioinf.cs.ucl.ac.uk/psipred/