
Статья опубликована в рамках: III Международной научно-практической конференции «Научное сообщество студентов: МЕЖДИСЦИПЛИНАРНЫЕ ИССЛЕДОВАНИЯ» (Россия, г. Новосибирск, 23 мая 2012 г.)
Наука: Биология
Секция: Экология
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов

ТОЛЕРАНТНОСТЬ ДРЕВЕСНЫХ РАСТЕНИЙ К ГИДРОТЕРМИЧЕСКОМУ РЕЖИМУ В РАЙОНЕ ИНТРОДУКЦИИ
Гриб Виктория Викторовна,
Левчук Марина Александровна
студенты 4 курса, кафедра экологии и рационального природопользования ФГБОУ ВПО «БГУ им. Ак. И. Г. Петровского», г. Брянск
Солдатова Валерия Викторовна
студентка 3 курса, кафедра биологии и экологии ФГБОУ ВПО Балашовского филиала (института) «СГУ им. Н. Г. Чернышевского»
E-mail: lubimov-v@mail.ru
Любимов Валерий Борисович
научный руководитель, доктор биол. наук, профессор, зав.кафедрой экологии и рационального природопользования ФГБОУ ВПО «БГУ им. ак. И. Г Петровского», г. Брянск
Актуальность исследований. Урбанизация, рост автопарка, развитие промышленности, гражданское строительство сопровождаются загрязнением окружающей среды, и ведет к ухудшению здоровья населения. Наиболее эффективным способом оздоровления окружающей среды в городах и поселках является создание насаждений различного целевого назначения, что определяет целесообразность интродукции древесных растений и разработка ассортимента из видов, характеризующихся высокой устойчивостью, декоративностью, другими хозяйственно-ценными качествами. Изучение толерантности растений к абиотическим факторам среды и их ответных реакций на силу воздействия факторов представляет большой, как научный, так и практический интерес. Такие исследования позволяют определить экологический спектр вида и лимитирующие интродукцию факторы и разработать приёмы, направленные на нейтрализацию факторов, сила которых выходит за пределы толерантности вида. Исследователями разработан ряд методов определения физиологических показателей водного режима, засухоустойчивости, солевыносливости и морозостойкости [1‑5, 8, 9, 12]. В первой половине двадцатого столетия была выдвинута гипотеза о возможности использования показателя жароустойчивости растений с целью отбора видов для южных районов страны, характеризующихся жесткими лесорастительными условиями. Например, метод, предложенный Ф. Ф. Мацковым, и в настоящее время используется для определения жароустойчивости растений при их интродукции в ту или иную природную зону [3‑7]. Этот метод включен в ряд учебных пособий и практикумов по физиологии растений [3‑5]. Однако изученность накопленного опыта указывает на весьма противоречивые результаты. Например, И. Д. Зелепухиным древесные растения Северной Америки, интродуцированные в Казахстан, разделены по степени жароустойчивости, методом Ф. Ф. Мацкова, на три группы: жароустойчивые — выдерживающие температуру +70 — +80°С, средней степени — +50— +60°С и нежароустойчивые — +45 — +48°С [8]. Согласно работам И. Д. Зелепухина жароустойчивость дуба черешчатого (Quercus robur L.), определенная методом Ф. Ф. Мацкова составляет +69°С, методом К. А. Ахматова — +58, методом В.П. Тарабрина — +51°С [1, 2, 8, 13]. Значительны и амплитуды летальных температур, определенные по разным методикам. Амплитуда летальных температур у видов древесных растений, определенная методом Ф. Ф. Мацкова, составляет 35°, методом К. А. Ахматова всего 7° и методом В.П. Тарабрина — 4°. Интерес представляют и результаты исследований по определению жароустойчивости растений, проведенные в пустыне — на полуострове Мангышлак. Исследователем методами В. П. Тарабрина, К. А. Ахматова и Ф. Ф. Мацкова была установлена толерантность 88 видов древесных растений из разных природных зон, в том числе и у представителей пустынь: саксаула черного (Haloxilon aphyllum (Minkw.) Jljin, c. белого (H. persicum Bge.), туранговых тополей (Turanga Bge.), видов из рода гребенщик (Tamarix L.), жузгуна (Calligonum L.) и др. В эксперименте были представители флор Европы, Средней, Центральной и Малой Азии, Дальнего Востока, Китая, Монголии, Японии и Северной Америки [10‑12]. Исследователь отмечает, что в не зависимости от условий обитания вида листья гибнут в температурном промежутке от +48 до +50°С. Как показывают фундаментальные исследования в области экологии и физиологии растений на Земном шаре высокие температуры в горячих источниках выдерживают лишь некоторые представители из отделов Бактерий и Сине-зеленых, или Циановых водорослей. Для остальной живой природы такая температура гибельна, тем более для растений, относящихся к группе пойкилотермных организмов, с температурой тела, зависящей от температуры окружающей Среды. Более того, наукой доказано, что все жизненно важные физиологические и биохимические процессы в зеленой клетке (дыхание, фотосинтез, движение цитоплазмы и др.) прекращаются в температурном промежутке от +48 до +50°С. При более высокой температуре клетка гибнет [6, 10, 12]. Однако в силу стереотипности мышления, несмотря на прогресс науки в этой области, продолжается интенсивный поиск «жароустойчивых» растений для их интродукции в аридные регионы, основанный на определении летальной температуры листьев в водяной бане. На основании изученности вопроса исследований следует сделать заключение о том, что для получения объективного результата по определению толерантности растений к высоким температурам необходимы комплексные, включая полевые и лабораторные, исследования с применением новых методов. В связи с этим нами были проведены экспериментальные исследования в Брянской и Саратовской области.
Цель исследования — оценить предложенные методы определения жароустойчивости растений и определить возможность их применения при определении перспективности вида для его интродукции в тот или иной регион. Выявить лимитирующие интродукцию факторы и разработать методы по оптимизации гидротермического режима содержания растений.
В задачу исследований входило изучение обоснованности термина жароустойчивости вида, базирующегося на летальных температурах листьев, а также причин получения в экспериментах столь противоречивых результатов, полученных разными методами при определении показателя жароустойчивости растений. Как известно, температура поверхности почвы в разные сезоны года и, даже, в течение суток значительно изменяется. Кроме того, температура поверхности почвы и приземного слоя отличается от температуры атмосферного воздуха. В связи с этим, мы ставили задачу, заключающуюся в решении вопроса, не является ли максимальная температура нагрева поверхности почвы и приземного слоя летальной для растений? При этом было намечено провести визуальные наблюдения за ювенильными растениями, выращиваемыми в открытом грунте и в лабораторных условиях на фоне погодных условий и выявить причину гибели проростков и ювенильных растений. Изучить гидротермический режим, определить абсолютную влажность завядания и разработать приёмы по оптимизации гидротермического режима содержания растений. Исследования осуществлялись студентами ФГБОУ ВПО «Балашовского филиала (института) Саратовского государственного университета им. Н. Г. Чернышевского» и ФГБОУ ВПО «Брянского государственного университета им. академика И. Г. Петровского».
Методы исследований – методами, разработанными Ф. Ф. Мацковым, К. А. Ахматовым и В. П. Тарабриным изучался показатель гибели листьев у разных видов, естественно обитающих в различных природных условиях и интродуцированных в Брянскую и Саратовскую область [2, 3, 9]. Сущность метода, предложенного Ф. Ф. Мацковым заключается в выдерживании срезанных листьев растений в водяной бане с температурой воды +40, +45 +50... +80°С, с последующим погружением листьев на 20 минут в слабый раствор соляной кислоты. Гибель листьев определяется по появлению бурого цвета на листовых пластинках. Температура, при которой появилась бурая окраска, соответствует (согласно методике) верхнему пределу жароустойчивости данного вида. По методике К. А. Ахматова при определении летальной температуры срезались побеги и вершинкой опускались в термосы наполненные водой разной температуры, а за тем побеги срезанной частью погружались в воду при комнатной температуре и проводились наблюдения за состоянием листьев и не одревесневших побегов. Температура водяной бани в термосе, которая вызвала гибель листьев и побегов является критической точкой за пределами, которой жизнь данного вида не возможна. По методике В. П. Тарабрина побеги не срезались, а наклонялись с растущих растений и верхушкой погружались в поднесенный бытовой термос, наполненный водой с определенной температурой. За опытными и контрольными побегами (контрольные побеги выдерживались в водяной бане при комнатной температуре) на растущих растениях проводились наблюдения, и устанавливалась температура гибели листьев и не одревесневшей части побега. Летальная температура определялась так же, как и в методике, описанной К. А. Ахматовым. Проведен также эксперимент по определению летальной температуры листьев у различных видов древесных растений методом, принципиально отличающимся от методов Ф. Ф. Мацкова, К. А. Ахматова и В. П. Тарабрина. Однолетние растения, выращенные в контейнерах, выдерживались в сушильном шкафу с горячим воздухом (+45, +4…+53°С). Выдержанные растения возвращались в питомник и в течение ряда дней за ними велись визуальные наблюдения, позволяющие определить температуру гибели листьев и не одревесневших побегов.
При изучении толерантности растений к гидротермическому режиму проведены экспериментальные исследования. Изучалась температура и влажность корнеобитаемого горизонта почвы в полевых и лабораторных условиях. В полевых условиях растения выращивались в открытом грунте, в лабораторных – в микро-посевных гидроизолированных чеках с постоянным подпитывающим через дренаж увлажнением. За состоянием растений проводились систематические наблюдения [12]. Определение абсолютной влажности почвы в периоды экстремальных температур и сухости атмосферного воздуха в Саратовской и Брянской области, а также абсолютной влажности завядания осуществлено по определенной методике [13, 14]. Результаты исследований обрабатывались методом математической статистики с применением компьютерной программы Microsoft Excel [15].
Результаты исследований. В результате собственных исследований и анализа литературных источников приходим к заключению, что независимо от экологических условий естественного обитания вида, летальная температура листьев, выдержанных в водяной бане составляет +49,3°С, ошибка среднего арифметического — 0,1°С, дисперсия — 1,27, критерий достоверности — 493, коэффициент вариации — 2,5 %, а показатель точности опыта — 0,2 %. Тенденции возрастания летальной температуры листьев по мере приближения вида (его естественного ареала) к пустынным регионам не наблюдается. Аналогичные результаты были получены студентами БФ(И)СГУ им. Н. Г. Чернышевского на кафедре биологии и экологии. В эксперимент были включены из числа голосеменных — лжетсуга Мензиса (Pseudotsuga menziesii (Mirb.) Franco), ель европейская (Picea excelsa Link.), ель колючая (Picea pungens Engelm.) и туя западная (Thuja occidentalis L.); из числа покрытосеменных — смородина черная (Ribes nigrum L.), береза бородавчатая (Betula verrucosa Ehm.), черемуха обыкновенная (Padus racemosa (Lam.) Gilib.), ива белая (Salix alba L.), тополь пирамидальный (Populus piramidalis Rosier) и из числа комнатных культур — традесканция и плющ. В первом опыте (метод Мацкова) летальная температура определялась у срезанных листьев по появлению бурого оттенка на листовой пластинке, во втором и третьем опыте (методы К. А. Ахматова и В. П. Тарабрина) по засыханию и скручиванию листьев и побегов, т.е. по истинной констатации гибели листьев и побегов. Как и следовало ожидать, по методике Ф. Ф. Мацкова визуально определить летальную температуру листьев оказалось невозможным, так как высокие температуры вызывают появление на листовых пластинках у разных видов различные цветовые оттенки (фиолетовые, темно-фиолетовые, бурые — различной степени насыщенности, оранжевые, желтоватые и зеленые цвета). Следует отметить, что дальнейшее повышение температуры не у всех видов сопровождается появлением бурого цвета, как описано в методике Ф. Ф. Мацкова, что и привело в работах многих исследователей к получению противоречивых результатов. В экспериментах, проведенных методами К. А. Ахматова и В. П. Тарабрина, получены четкие идентичные результаты. Листья всех, включенных в эксперимент видов, в независимости от условий их естественного местообитания, погибли при температуре близкой к + 50°С. Аналогичные результаты были получены и при выдерживании растений в сушильном шкафу.
В опытах по изучению гидротермического режима и его оптимизации для ювенильных растений получены следующие результаты. Исследования показали, что температура на поверхности почвы значительно ниже температуры атмосферного воздуха и не представляет угрозы для растений. Опыт проведен в: Аркадакском районе Саратовской области 15 августа 2010 г. в 15 часов. Температура атмосферного воздуха на высоте груди составляла +![]() С (на поверхности почвы -
С (на поверхности почвы - ![]() С), в Балашовском районе (экспериментальный участок БФ(И)СГУ им. Н. Г. Чернышевского) 16 августа 2010 г. в 15 часов, температура атмосферного воздуха на высоте груди составляла +
С), в Балашовском районе (экспериментальный участок БФ(И)СГУ им. Н. Г. Чернышевского) 16 августа 2010 г. в 15 часов, температура атмосферного воздуха на высоте груди составляла +![]() С (на поверхности почвы -
С (на поверхности почвы -![]() , а также в г. Брянске (экспериментальный участок БГУ им. ак. И. Г. Петровского) 15 августа 2010 г. в 16 часов, температура атмосферного воздуха на высоте груди составляла +
, а также в г. Брянске (экспериментальный участок БГУ им. ак. И. Г. Петровского) 15 августа 2010 г. в 16 часов, температура атмосферного воздуха на высоте груди составляла +![]() (на поверхности почвы -
(на поверхности почвы - ![]() , на глубине 20 см —
, на глубине 20 см — 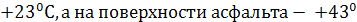 С). Таким образом, в самый экстремальный период (июнь – август, 2010 г.) температура на поверхности почвы и приземного слоя атмосферного воздуха была значительно ниже летальной температуры для растений (+
С). Таким образом, в самый экстремальный период (июнь – август, 2010 г.) температура на поверхности почвы и приземного слоя атмосферного воздуха была значительно ниже летальной температуры для растений (+![]() . Кроме температуры изучена и абсолютная влажность почвы. Результаты исследований отражены в табл. 1.
. Кроме температуры изучена и абсолютная влажность почвы. Результаты исследований отражены в табл. 1.
Таблица 1
Абсолютная влажность почвы (август 2010 г.)
Глубина взятия образца, см |
Масса сырой почвы, г |
Абсолютно сухая почва, г |
Кол-воды в образце, г |
Абсолютная влажность почвы, %
W= |
Аркадакский район Саратовской области - 18 августа 2010 г.
0-5 |
224 |
210 |
14 |
6,7 |
10-15 |
238 |
219 |
19 |
8,6 |
20-25 |
209 |
189 |
20 |
10,6 |
Балашовский район Саратовской области 16 августа 2010 г.
0-5 |
192 |
180 |
12 |
6,7 |
10-15 |
204 |
190 |
14 |
7,4 |
20-25 |
208 |
186 |
22 |
11,8 |
Брянск - экспериментальный участок Брянского государственного университета имени академика И.Г. Петровского- 15 августа 2010 г.
0-5 |
186,4 |
174,14 |
12,26 |
7,0 |
10-15 |
185,8 |
169,5 |
16,3 |
9,6 |
20-25 |
186,0 |
164,0 |
22,0 |
13,4 |
w — абсолютная влажность почвы, а –кол-во воды в образце, р – масса абсолютно сухой почвы.
C глубиной абсолютная влажность почвы повышается с 6,7 % в поверхностном горизонте до 11,8 % на глубине 20-25 см (г. Балашов) и, соответственно, с 7,0 % до 13,4 % (г. Брянск). Здесь следует отметить, что у многих 1‑2-летних древесных растений корневые системы уходят в более глубокие слои почвы, что обеспечивает сохранение оптимального водного баланса. Приведем годичный прирост за 2010 год у ряда интродуцентов в г. Брянске и г. Балашове (табл.2).
Таблица 2
Высота 1-2 летних растений, в см (15 августа 2010 г.)
Вид |
M±m, см |
|
|
|
t |
P |
N |
г. Брянск
Quercus rubra L.-1 лет |
11,26±0,2 |
1,87 |
1,37 |
12,1 |
46,5 |
2,15 |
32 |
Q. rubra L.-2 лет |
25,9±1,17 |
44,4 |
6,66 |
25,6 |
22,1 |
4,53 |
32 |
Forsythia europaea Deg. et. Bald.-1 лет |
111,9±2,9 |
271,1 |
16,5 |
14,7 |
38,5 |
2,59 |
32 |
г. Балашов
Вид |
M±m, см |
|
|
|
t |
P |
N |
Q. rubra L.-1 лет |
8,9±0,19 |
1,18 |
1,1 |
12,2 |
46,4 |
2,16 |
32 |
Q. rubra L.-2 лет |
20,9±0,48 |
7,44 |
2,73 |
13,05 |
43 |
2,3 |
32 |
Mahonia aquifolium (Pursh) Nutt.- 2 лет |
18,87±0,3 |
2,69 |
1,64 |
13,82 |
40,9 |
2,44 |
32 |
В результате исследований получены основные параметры, дающие довольно полное представление о средних арифметических значениях и заслуживающие доверия на 95 % доверительном уровне, ввиду большой достоверности средних арифметических (t>3) и значений показателя точности опыта меньшего 5 % (Р<5%). Следует отметить, что, несмотря на экстремальные погодные условия, рост и развитие растений в середине августа 2010 г. продолжался. Вегетирующие побеги и листья у включенных в эксперимент видов небыли повреждены высокими температурами, и падения тургора не наблюдалось даже в дневные часы. Абсолютная влажность почвы в питомнике г. Балашова составила в корнеобитаемом горизонте 11,8 %, а в Брянске – 13,4 %. Сравнение средних высот одно и двухлетних растений Quercus rubra L. показало на достоверность превышения растений, выращиваемых в Брянске, над растениями, выращиваемыми в г. Балашове. Табличное значение критерия Стьюдента для наших вариационных рядов t=2, что значительно ниже вычисленных значений. Следовательно, средние арифметические значения, полученные для высот растений в г. Брянске достоверно превышают средние арифметические значения, полученные для высот растений, выращиваемых в г. Балашове, что определяется разностью влажности корнеобитаемого горизонта почвы. Однако в периоды экстремальных температур поверхность почвы в южных районах (сухие степи, полупустыни, пустыни) может нагреваться до пятидесяти градусов и выше, что вызывает массовую гибель всходов и растений ювенильного возраста, но это происходит только в случае острого дефицита влаги в почве. При наличии оптимального для растений увлажнения (не ниже 75 % от полевой влагоемкости), высокие температуры способствуют интенсивному росту растений.
В августе 2010 г. был проведен лабораторный опыт по определению влажности завядания (ВЗ) методом вегетационных миниатюр для проростков пузыреплодника калинолистного - Physocarpus opulifolius (L.) Maxim. и клена остролистного -Acer platanoides L. Результаты отражены в табл. 3.
Таблица 3
Определение абсолютной влажности почвы (ВП) и абсолютной влажности завядания (ВЗ) для проростков Physocarpus opulifolius (L.) Maxim. Балашов, 2010 г. (посев 3 августа 2010 г.)![]()
Дата |
19.8 |
20.8 |
21.8 |
22.8 |
23.8 |
24.8 |
25.8 |
26.8 |
|
32 |
30 |
29 |
29 |
27 |
27 |
27 |
26 |
Масса образца почвы, г |
174,8 |
164,9 |
162,5 |
152,7 |
138,5 |
115,6 |
100,8 |
85,5 |
Масса воды, г |
98,2 |
88,3 |
85,9 |
76,1 |
61,9 |
39,0 |
24,2 |
8,9 |
Абсолютная ВП, % |
128 |
115 |
112 |
99 |
80 |
50,9 |
31,0 |
11,6 |
Абсолютно сухая почва составила 76,6 г. 26 августа зарегистрировано интенсивное падение тургора, что оценено нами, как ВЗ. 27 августа все проростки погибли. Абсолютная влажность завядания для проростков составляет 11,6 %. Аналогичные результаты нами получены и в условиях г. Брянска. Абсолютная влажность почвы (%) определялась по формуле: W=![]() ; а –кол-во воды в образце, р – масса абсолютно сухой почвы. При этом следует отметить, что семена Physocarpus opulifolius (L.) Maxim. и других видов проросли, а всходы успешно развивались при высоких температурах, которые были выше, чем температуры, зарегистрированные на поверхности почвы в самые жаркие периоды 2010 г. Анализ опыта подтверждает, что для растений в периоды высоких температур атмосферного воздуха губительны не температуры, а дефицит влаги. При размножении растений, безусловно, необходимо анализировать экологический спектр вида, выявлять экологические факторы, лимитирующие его интродукцию и моделировать условия, обеспечивающие, прежде всего, оптимальный гидротермический режим, соответствующий условиям его естественного обитания. В условиях стабильного, регулярного обеспечения водой у растений, в результате транспирации, температура листьев близка к температуре окружающей среды и значительно ниже температуры, приводящей к летальному исходу. Однако в периоды экстремальных температур, сухости воздуха и почв, при отсутствии регулярного орошения, нарушение водного баланса у растений приводит к нагреванию листьев до летального исхода, что часто наблюдается в насаждениях южной и юго-восточной части территории Саратовской области, относящейся к сухой степи и граничащей с полупустыней.
; а –кол-во воды в образце, р – масса абсолютно сухой почвы. При этом следует отметить, что семена Physocarpus opulifolius (L.) Maxim. и других видов проросли, а всходы успешно развивались при высоких температурах, которые были выше, чем температуры, зарегистрированные на поверхности почвы в самые жаркие периоды 2010 г. Анализ опыта подтверждает, что для растений в периоды высоких температур атмосферного воздуха губительны не температуры, а дефицит влаги. При размножении растений, безусловно, необходимо анализировать экологический спектр вида, выявлять экологические факторы, лимитирующие его интродукцию и моделировать условия, обеспечивающие, прежде всего, оптимальный гидротермический режим, соответствующий условиям его естественного обитания. В условиях стабильного, регулярного обеспечения водой у растений, в результате транспирации, температура листьев близка к температуре окружающей среды и значительно ниже температуры, приводящей к летальному исходу. Однако в периоды экстремальных температур, сухости воздуха и почв, при отсутствии регулярного орошения, нарушение водного баланса у растений приводит к нагреванию листьев до летального исхода, что часто наблюдается в насаждениях южной и юго-восточной части территории Саратовской области, относящейся к сухой степи и граничащей с полупустыней.
В результате экспериментальных исследований мы получили подтверждение выдвинутой гипотезы, которая заключается в том, что в процессе эволюции у растений выработались механизмы гомеостаза, обеспечивающие их равновесное состояние при изменении положительных температур не путем повышения жароустойчивости клетки, а путем регуляции водообмена, позволяющего поддерживать температуру клеток, не превышающую температуру окружающей среды. Гипотеза находит подтверждение и при анализе формирования ландшафтов Земли. Безусловно, что температура (но минимальная) - это показатель, ограничивающий распространение видов на Земле. Среди растений имеются как эвритермные, так и стенотермные виды. В таблице 4 приведена экологическая валентность растений по отношению к температурному фактору, эволюционно сформировавшаяся в конкретных регионах земного шара. Амплитуда максимальных температур воздуха в пределах произрастания древесных растений составляет всего 10‑15 градусов.
Таблица 4
Экологическая валентность растений по отношению к температурному фактору
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
В условиях пустынь и полупустынь, в процессе эволюции, большинство растений приспособились переносить высокие температуры не повышением устойчивости зеленой клетки, а завершением вегетационного периода до наступления высоких температур и, как следствие, засухи, т.е. до выхода силы влияния этого экологического фактора за пределы толерантности данной группы видов растений. Это - эфемеры и эфемероиды. Первые - переносят длительный неблагоприятный период в виде семян, вторые - подземными органами: луковицами, клубнями и корневищами. Поверхность почвы нагревается в пустынях до 60 и более градусов (табл. 5). На высоте двух метров от поверхности почвы температура на 15‑28 градусов ниже и, следовательно, зеленая клетка древесных растений не испытывает прямого воздействия летальных для нее температур. Однако семенное возобновление древесных растений пустынь и полупустынь часто гибнет под воздействием высоких (более +45°С) температур поверхности почвы и быстрого иссушения ее верхних горизонтов. И только в отдельные, благоприятные для роста и развития растений, годы сохраняется естественное возобновление. Учитывая, что летальная температура листьев древесных растений превышает максимальную температуру воздуха даже пустынь, можно сделать вполне обоснованное заключение о том, что высокая температура воздуха не является основным лимитирующим интродукцию фактором.
Таблица 5
Зависимость температуры атмосферного воздуха от высоты над уровнем почвы (полуостров Мангышлак, 1989 г.)
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
За пределы экологической валентности выходят не максимальные температуры воздуха, а, чаще всего, минимальные температуры и (практически всегда) – дефицит влаги, о чем свидетельствует накопленный большой фактический материал по переселению растений в разные регионы земного шара [11‑12, 14].
Амплитуда минимальных температур для разных регионов, в пределах произрастания древесных растений, составляет около 105°С (табл.4). Безусловно, что в биоценозах, при положительных температурах (бассейны рек Конго и Амазонки) сформировались стенотермные растения. А виды, произрастающие в сухих субтропиках, северных и южных пустынях, в условиях степи, лесостепи и лесной зоне и севернее – эвритермные. Это объясняется, прежде всего, аксиомой Ч. Дарвина об адаптивности вида к абиотическим факторам среды обитания и экологическими законами формирования в процессе эволюции экологического спектра вида и границ его современного ареала. В таблице 6 приведены усредненные гидротермические условия физико-географических поясов земного шара, т.е. условия формирования биоценозов. Изученность накопленного фактического материала показывает, что важнейшими экологическими факторами, формирующими (в процессе эволюции) природные экосистемы является радиационный баланс и валовое увлажнение. Если рассматривать эти факторы в пределах одного физико-географического пояса Земли, то нетрудно убедиться в том, что амплитуда гидротермического коэффициента (ГТК) в результате которого
Таблица 6
Гидротермический коэффициент физико-географических поясов земного шара
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
сформировались различные ландшафты, например, от пустынь, полупустынь, степей до смешанных лесов и тайги, лесотундры и тундры в большей степени зависит от показателя валового увлажнения. Приведем гидротермические показатели ландшафтов (Табл. 7).
Таблица 7
ГТК основных типов ландшафтов (по А.М. Рябчикову)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Таким образом, основными, лимитирующими видовое разнообразие фитоценозов факторами являются отрицательные температуры и валовое увлажнение, что подтверждает нашу гипотезу.
Научная новизна и практическая значимость. Установлено, что лимитирующим интродукцию фактором являются не максимальная температура в районе исследований, а абсолютный минимум и дефицит воды. Определен оптимальный гидротермический режим для размножения и содержания интродуцентов, что может быть использовано при выращивании посадочного материала и создания насаждений.
Достоверность исследований обеспечена большим объемом собранного в результате многолетних исследований экспериментального материала с применением современных методов в области экологии и его обработкой методом математической статистики, с использованием компьютерной программы Microsoft Excel 3.
Заключение. При определении летальной температуры разница для различных видов столь незначительна, что дифференцировать виды на ее основании, по степени жароустойчивости, не представляется возможным. Температура гибели листьев (+ 50°С) значительно выше температуры атмосферного воздуха даже в условиях пустынь. Высокие температуры атмосферного воздуха при достаточном водообеспечении не только не губительны, а способствуют повышению энергии роста, продуктивности и не являются лимитирующим интродукцию фактором. К высоким температурам атмосферного воздуха и его сухости растения адаптировались путем эволюции механизмов гомеостаза, обеспечивающих равновесное состояние организмов в изменяющихся условиях среды, и, прежде всего, обеспечивающих оптимальный водный режим. Циркуляция воды (почва – растение - атмосфера) позволяет поддерживать температуру зеленой клетки на уровне не выше температуры окружающей среды. Поиск перспективных интродуцентов следует вести не по показателю гибели листьев от высокой температуры, а по степени приспособленности или толерантности растений к дефициту влаги и минимальным температурам.
Список литературы:
1.Андрейченко Л. М. Интродукция дуба в Северную Киргизию / Л. М. Андрейченко. - Фрунзе: Илим, 1978.‑ С. 118.
2.Ахматов К А. Адаптация древесных растений к засухе / К.А. Ахматов. - Фрунзе: Илим, 1976.‑200 с.
3.Викторов Д. П. Малый практикум по физиологии растений.- М.: Высшая
школа, 1983.‑136 с.
4.Генкель П. А. Физиология растении / П. А. Генкель. - Л.-М.: Просвещение, 1975.‑336 с.
5.Генкель П. А. 0 состоянии и направлении работ по физиологии жаро- и засухоустойчивости растений // Проблемы засухоустойчивости растений.- М.: Наука, 1978.-С. 5‑20.
6.Горышина Т. К. Экология растений / Т. К. Горышина. - М.: Высшая школа, 1975.‑368 с.
7.Гурина Т. Ф. Сравнительная оценка жароустойчивости интродуцированных хвойных экзотов / Т. Ф. Гурина.- Пущино, 1984.‑ С. 50.
8.Зелепухин И. Д., Дурова Р. А. Жароустойчивость североамериканских деревьев и кустарников в Алма-Ате //Труды бот. сада АН Каз ССР.- Алма-Ата: Наука. - Т. 11 .‑С. 113.
9.Тарабрин В. П. Жароустойчивость древесных растений и методы ее определения / В. П. Тарабрин // Бюл. ГБС, 1969.- Вып.74.‑ С. 53‑56.
10.Любимов В. Б. К вопросу о методах определения жароустойчивости древесных растений / В. Б. Любимов.- Шевченко: МЦНТИ, 1988.‑ 4 с.
11.Любимов В. Б. К вопросу о жароустойчивости растений / В. Б. Любимов, Н. П. Котова, Р. Н. Ломадзе. - Волгоград: Вестник Вол. ГУ. Серия 3. «Экономика. Экология», № 2.- 2009. – С. 238‑244.
12.Любимов В. Б. Толерантность растений к дефициту влаги и температурному режиму / В. Б. Любимов, Н. П. Котова. - Брянск: Вестник РИО БГУ, 2010. – С. 185‑190.
13.Любимов В. Б. Методика изучения в полевых условиях третьей среды обитания живых организмов – почвы // В. Б. Любимов, Л. Г. Горковенко, Е. Б. Смирнова. – Балашов: БФСГУ, 1998. – 13 с.
14.Любимов В. Б. Интродукция растений (теория и практика) / В. Б. Любимов. – Брянск: Курсив, 2009. – 365 с.
15.Любимов В. Б. Математические методы и их практическое применение в экологических исследованиях / В. Б. Любимов, К. В. Балина. – Брянск: РИО БГУ, 2010. – 176 с.
дипломов