
Статья опубликована в рамках: XXXVII Международной научно-практической конференции «Научное сообщество студентов XXI столетия. ЕСТЕСТВЕННЫЕ НАУКИ» (Россия, г. Новосибирск, 14 января 2016 г.)
Наука: Биология
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
отправлен участнику

ОСОБЕННОСТИ КЛЕТОК КОРНЕЙ ТРАНСГЕННЫХ РАСТЕНИЙ TАБАКА СО ВСТРОЕННЫМ АГРОБАКТЕРИАЛЬНЫМ ГЕНОМ iaaM
Лаврентьева Вероника Валерьевна
студент 1 курса магистратуры, кафедра физиологии и биохимии растений УрФУ, г. Екатеринбург
Е-mail: lavrentieva.w@gmail.com
Киселева Ирина Сергеевна
научный руководитель, кандидат биологических наук, доцент, зав.кафедрой физиологии и биохимии растений УрФУ, г. Екатеринбург
В настоящее время в качестве моделей для изучения механизмов действия гормонов растений часто используют трансгенные растения с измененным уровнем экспрессии генов синтеза фитогормонов. Такие модели позволяют исследовать все уровни действия гормонов – от организменного до молекулярного.
Ауксины являются одними из важнейших гормонов растений, обладают множественными физиологическими эффектами. Имея трансгенное растение со встроенными генами синтеза ауксинов, например, агробактериальными генами iaaM, исследователи могут изучать ауксин зависимые процессы и их роль в жизнедеятельности растительного организма.
Целью данной работы была оценка влияния экспрессии агробактериального гена биосинтеза ауксинов iaaM на морфологию, анатомию и рост корней табака. В литературе имеются некоторые сведения о таких трансгенных растениях, главным образом, об особенностях строения побегов и физиолого-биохимических признаках листьев, однако публикаций, посвященных изучению корней мало [1, 5, 6, 7], хотя известно, что ауксины обладают эффектом стимуляции ризогенеза.
Объекты и методы исследования
Объектом исследования были черенки растений табака сорта Самсун: контрольные (нетрансформированные), трансгенный контроль (растения, трансформированные «пустой» плазмидой pSS) и трансгенные, содержащие генетическую конструкцию с целевым агробактериальным геном iaaM под сильным конститутивным промотором 35S. Растения были получены в Лаборатории биотехнологии растений ФИБХ РАН и любезно предоставлены для работы Е. Б. Рукавцовой, В. В. Алексеевой и А. А. Ермошиным.
В работе использовали придаточные корни черенков, которые срезали с выращенных в почве растений табака и помещали в широкие пробирки в воду. Каждый черенок имел 8-10 узлов и 5-10 листьев. Всего в работе было использовано 10 растений контрольной линии, 4 растения линии pSS, 8 трансгенных растений линии 6/1 и 26 растений линии 6/2. По данным Алексеевой В.А. [1], исходные растения этих линий имели различный фенотип: растения линии 6/2 были ниже, имели более сильно скрученные листья в сравнении с растениями линии 6/1. То есть, линия 6/2 имела аномальный фенотип.
Придаточные корни по мере достижения ими длины 1,5-2 см фиксировали в уксуснокислом алкоголе. Готовили давленые препараты меристематической части корней и продольные срезы корней в зоне растяжения [3, 4]. Препараты окрашивали ацетолакмоидом или ацетокармином. При помощи окуляр-микрометра и объект-микрометра определяли линейные размеры длинной и короткой оси клетки, рассчитывали объем клеток [2] и индекс прозенхимности, за который принимали отношение длины клетки к ее ширине.
Статистическую обработку данных проводили в пакете прикладных программ «STATISTICA 6» (Statsoft). В каждом случае с одного побега фиксировали 3-5 корешков. В каждом из них определяли размеры 25-30 клеток соответствующей зоны. Все значения измеряемых или расчетных параметров усредняли в пределах каждого побега. Индивидуальный побег принимали за биологическую повторность. На рисунках результаты представлены в виде среднего значения и стандартной ошибки среднего.
Результаты и обсуждение
Увеличение корня в длину происходит за счет активного деления клеток КАМ и растяжения клеток в зоне роста. Известно, что ауксины регулируют растяжение клеток. В связи с этим мы предположили, что объем и индекс прозенхимности клеток корня трансгенных растений табака со встроенным геном iaaM будет больше, чем у контрольных растений. Результаты приведены на рисунках 1 и 2.
По нашим наблюдениям во всех вариантах клетки меристемы имели типичную для зоны деления кубическую форму. Объем клеток во всех линиях существенно не отличался и составлял от 2000 до 2750 мкм3. Индекс прозенхимности у растений контрольной линии, трансгенного контроля и трансгенной линии 6/1 не имел достоверных отличий и составлял 1,04 – 1,05.
Таким образом, меристематические клетки трансгенных растений с повышенным уровнем ауксинов имели ту же форму и размеры, что и у контрольных растений. Следовательно, избыток ИУК в зоне деления не оказывал влияния на морфологические характеристики клеток.



Рисунок 1. Диаметр, объем и индекс прозенхимности клеток меристемы корней растений табака (среднее значение ± ошибка среднего)
Дифференцированные клетки корня имели вытянутую цилиндрическую форму без выростов. Существенных отличий по длине большой оси клеток не наблюдали, в то время как длина короткой оси (ширина) клеток у контрольных растений была заметно больше. Объемы клеток варьировали от 92000 до 130000 мкм3. Достоверно отличалась от остальных только линия трансгенного контроля. Индекс прозенхимности у «ауксиновых» растений был достоверно выше, чем у растений контрольной линии.
Это позволяет сделать вывод о том, что линии с избыточной продукцией ауксина по сравнению с контрольными линиями растягиваются в большей степени вдоль длинной оси клетки и в меньшей степени – вдоль короткой.
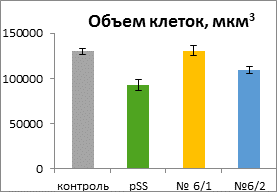


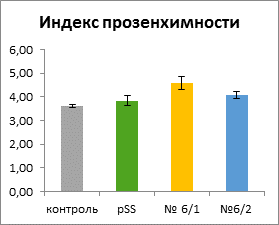
Рисунок 2. Длина, ширина, объем клеток и индекс прозенхимности в зоне растяжения корней табака всех линий (среднее значение ± ошибка среднего)
В зоне растяжения корня избыток экзогенной ИУК не вызывал изменения объема клеток у линии 6/1 с нормальным фенотипом, но приводил к уменьшению объема клеток линии 6/2 с аномальным фенотипом, и при этом обусловливал формирование более узких и длинных клеток. Можно предположить, что в условиях повышенных концентраций ИУК в клетках трансгенных растений происходило изменение ориентации элементов цитоскелета, определяющих направление роста клеток.
Как было отмечено выше, по данным Алексеевой В.А. [1], «ауксиновые» растения имели различный фенотип. Вероятно, у линии 6/2, имеющий аномальный фенотип, в большей степени, чем у линии 6/1 был нарушен баланс гормонов, что проявилось не только в нетипичном габитусе, но и в изменении формы и размеров клеток. У растений трансгенного контроля клетки корня в зоне растяжения были мельче, чем у нетрансгенного контроля и у «ауксиновых» растений. Поскольку инсерция генетической конструкции носит случайный характер, можно предположить, что она встроилась в функционально активный участок генома растения, что привело к некоторым нарушениям, проявляющимся в торможении роста.
Заключение
Тканевая организация придаточных корней контрольных и трансгенных растений не отличается: в молодых корнях имеется 2-3 слоя клеток коры, эндодерма, центральный цилиндр. Не обнаружены отличия в форме и размерах клеток меристемы корня у трансгенных и контрольных растений. В зоне растяжения корня наблюдали увеличение индекса прозенхимности клеток у «ауксиновых» растений в сравнении с контрольными, при этом объемы клеток существенно не отличались.
Список литературы:
-
Алексеева В. В., Рукавцова Е. Б., Бобрешова М. Е., Ложникова В. Н.,
Бурьянов Я. И. Получение и анализ трансгенных растений табака, экспрессирующих агробактериальный ген триптофамонооксигеназы // Физиология растений. – 2004. –Т. 51, №4. – С. 600-606. - Борзенкова Р. А, Храмцова Е. В. Определение мезоструктурных характеристик фотосинтетического аппарата растений. – Екатеринбург: Издательство УрГУ им. А.М. Горького, 2006. - 27 с.
- Паушева З. П. Практикум по цитологии растений. – Москва: Агропромиздат, 1988. – 271 стр.
- Прохорова И. М., Ковалева М. И., Фомичева А. Н. Оценка митотоксического и мутагенного действия факторов окружающей среды. – Ярославль: Ярославский государственный университет, 2003. - 5 с.
- Eklöf S, Astot C, Sitbon F, Moritz T, Olsson O, Sandberg G . Transgenic tobacco plants co-expressing Agrobacterium iaa and ipt genes have wild-type hormone levels but display both auxin- and cytokinin-overproducing phenotypes //The Plant Journal. – 2000. – V.23, N 2. – P. 279-284.
- Sitbon F., Sundberg B., Olsson O., and Sandberg G. Free and Conjugated Indoleacetic Acid (IAA) Contents in Transgenic Tobacco Plants Expressing the iaaM and iaaH IAA Biosynthesis Genes from Agrobacterium tumefaciens // Plant Physioiogy. – 1991. – V. 95, № 2. – P. 480–485.
- Sitbon, F., Hennion, S., Sundberg, B., Little, C.H.A., Olsson, O., Sandberg, G. 1992a. Transgenic tobacco plants coexpressing Agrobacterium tumefaciens iaaM and iaaH genes display altered growth and indoleacetic acid metabolism. Plant Physiol. 99: 1062–1069.
отправлен участнику