
Статья опубликована в рамках: XXXII Международной научно-практической конференции «Научное сообщество студентов XXI столетия. ЕСТЕСТВЕННЫЕ НАУКИ» (Россия, г. Новосибирск, 09 июня 2015 г.)
Наука: Медицина
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
отправлен участнику

БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ОБРАЗОВАНИЯ СИНАПСОВ
Ишкинин Руслан Эдуардович
студент 4 курса, Башкирский государственный медицинский университет, РФ, Республика Башкортостан, г. Уфа
E -mail: ruslan.9031993@yandex.ru
Насртдинов Ильмир Галинурович
студент 4 курса, Башкирский государственный медицинский университет, РФ, Республика Башкортостан, г. Уфа
E -mail: i.nasrtdinov@mail.ru
Султанов Руслан Айратович
студент 4 курса, Башкирский государственный медицинский университет, РФ, Республика Башкортостан, г. Уфа
E -mail: sultanovruslan@mail.ru
Князева Ольга Александровна
научный руководитель, д-р биол. наук, профессор кафедры биологической химии, Башкирский государственный медицинский университет, РФ, Республика Башкортостан, г. Уфа
Современные представления о молекулярных механизмах формирования долговременной памяти основаны на концепции, согласно которой приобретение нового опыта сопровождается экспрессией широкого спектра новых белков с последующими структурными изменениями синаптических контактов между клетками [2]. Каким же образом образуются новые синаптические связи, играющие одну из ключевых ролей в формировании долговременной памяти? На основании уже известных фактов нами предложена гипотеза, которая может раскрыть один из аспектов этого сложного явления. Для ее обоснования были использованы следующие данные:
1. В экспериментальных исследованиях на крысах было показано, что количество синапсов увеличивается с приобретением опыта [4].
2. Движение медиаторов в нейроно-глиальных щелях влияет на локализацию нейрорецепторов. Рецепторы, вытесненные из синаптической щели, кластеризуются у квазистационарных точек, образуя на нейроне мозаику метаботропных рецептивных кластеров (МРК).
3. Медиаторы, интерферирующие за пределами синапсов, управляют локальными конформациями МРК [2].
4. При введении серотонина больным шизофренией отмечается улучшение памяти и внимания [1].
5. Изменения активности серотонинергической системы приводит к нарушению синаптической пластичности, дисбалансу нейрональных связей и, как следствие, развитие нейродегенеративных заболеваний.
6. Рецепторы серотонина являются как метаботропными (5-НТ 1,2,4,5,6,7), так и ионотропными (5-НТ 3). Причем 5-НТ 1,5 снижает клеточный уровень цАМФ, 5-НТ 4, 6, 7 — увеличивает его. 5-НТ 2 рецепторы повышают клеточный уровень IP3 и DAG [3].
7. Агонисты 5-НТ 2А рецепторов приводят к улучшению настроения, физической и психической активности, улучшению ассоциативного и креативного мышления.
8. Мозаприд (агонист 5-НТ 4) способствует нейрогенезу в ЖКТ [6].
9. СИОЗС устраняет когнитивные дисфункции, вызываемые сахарным диабетом.
10.Формирование у крыс пищевых условных рефлексов сопровождается повышением содержания РНК в ядре и цитоплазме [5].
11.При формировании условных рефлексов увеличивается степень метилирования ДНК мозга. Гиперметилирование CpG-островков в промоторных областях генов, приводит к устойчивой репрессии транскрипции. Репрессия транскрипции в этом случае опосредована метилцитозин-связывающими белками, которые способны связываться с метилированными CpG-динуклеотидами.
12.Ингибирование белка РР1 приводит к уменьшению проблем с памятью.
13.CaM-киназа фосфорилирует CREB (цАМФ-зависимый транскрипционный фактор) в течение нескольких секунд после притока Ca2+.
14.К генам, транскрипция которых регулируется CREB, относятся c-fos, BDNF, NGF, VGF (белки, участвующие в процессах памяти).
15.Доказана роль CREB в нейрональной пластичности и долговременной памяти.
16.Фактор роста нервов (NGF) вызывает рост аксонов: способствует их ветвлению и небольшому удлинению. NGF связывается, по меньшей мере, c двумя классами рецепторов: LNGFR и TrkA.
17.Трофические эффекты NGF могут передаваться на развивающиеся нейроны в виде ретроградного транспорта NGF от нервных терминалей в тела клеток. NGF активно захватывается в нервные терминали и транспортируется ретроградно в сому.
18.Если NGF вводится взрослым крысам, у них улучшается выполнение пространственных задач [3].
19.VGF-белок, участвующий в регулировании энергетического гомеостаза, метаболизма и синаптической пластичности. Экспрессия белка VGF индуцируется нейротрофными факторами NGF, BDNF и нейротрофином-3. Сниженная экспрессия VGF отмечается при болезни Альцгеймера, с развитием которой происходит потеря долговременной памяти [6].
Исходя из вышеперечисленного, можно предположить, что при прохождении импульса по аксону к терминали в синаптическую щель, выделяется нейромедиатор, который взаимодействует с постсинаптической мембраной и частично вымывается в нейроглиальную щель. Там он взаимодействует с метаботропными рецепторами, расположенными на соседних нейронах. Под действием медиатора происходят конформационные изменения рецептора, что увеличивает чувствительность к медиатору (возможно, эти изменения лежат в основе образования временной связи между группами нейронов при кратковременной памяти).
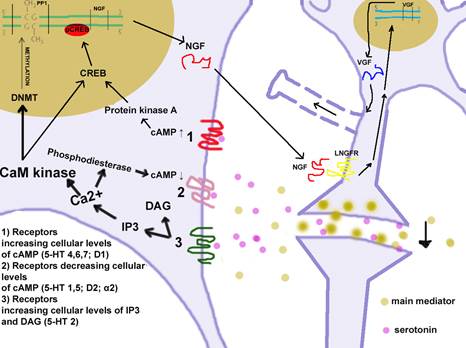
Рисунок 1. Образование синаптических связей
Метаботропный рецептор передает сигнал внутрь клетки и многократно его усиливает. Эти рецепторы связаны с G-белком, которые могут увеличивать или уменьшать концентрацию вторичных мессенджеров (цАМФ, инозитол-3-фосфата (ИТФ), диацилглицерола (ДАГ)). При увеличении внутриклеточного уровня цАМФ активируется протеинкиназа А, которая перемещается в клеточное ядро, где фосфорилирует и тем самым активирует CREB. Активированный CREB связывается с CRE-участком промотора, при этом начинают транскрибироваться гены BDNF, NGF. При уменьшении уровня цАМФ протеинкиназа А становится неактивной, не фосфорилирует СREB и не запускает синтез белков. При увеличении содержания ИТФ увеличивается концентрация эндогенного Ca2+, а ДАГ — экзогенного. ДАГ и Ca2+ активируют CaM-киназу. Она фосфорилирует CREB, который активирует синтез нейротрофинов. СaM-киназа также активирует ДНК-метилтрансферазу.
Метилирование ДНК заключается в присоединении метильной группы к цитозину в составе CpG-динуклеотида в позиции С5 цитозинового кольца. Гиперметилирование CpG-островков в промоторных областях генов, приводит к устойчивой репрессии транскрипции. К белкам репрессорам относятся метилцитозин-связывающие белки. Возможно к ним также относится белок РР1. Так как содержание РНК увеличивается, значительная часть генов растормаживается. Поэтому можно предположить, что кодируемые активированными генами белки ответственны за формирование синапсов. Ca2+ также активирует фосфодиэстеразу, которая расщепляет цАМФ до АМФ. Это приводит к тому, что активация синтеза нейротрофинов идет только по одному пути. NGF выходит в нейроглиальную щель и связывается c двумя классами рецепторов: LNGFR и TrkA. NGF активно захватывается в нервные терминали и транспортируется ретроградно в сому. Там он действует на экспрессию белка VGF и др. белков участвующих в образовании синапса. Белок VGF участвует в регулировании энергетического гомеостаза, метаболизма и синаптической пластичности.
Таким образом, мембрана нейрона начинает расти к месту секреции NGF. При сближении двух мембран между ними образуется синапс, образование которого следует считать завершением формирования долговременной памяти (рис. 1).
Список литературы:
1.Никольский Е.Е. Молекулярные механизмы передачи информации через синапсы химического типа // Казан. мед. журн. — 2010. — Т. 91, — № 4. — С. 433—437.
2.Радченко А.Н. Информационный ключ к памяти мозга // Труды СПИИРАН. Вып. 3, т. 2. СПб: Наука, 2006. — С. 269—286.
3.Скребицкий В.Г. Синаптическая пластичность как проблема нейрофизиологии. //Вестник РФФИ. — 2004. — Т.4. — С. 65—81.
4.Тушмалова Н.А. Современные представления о макромолекулярных механизмах памяти//Исследование памяти / Под ред. Н.Н. Корж. М., 2009. — С. 137—145.
5.Bushey D., Tononi G., Cirelli C. Sleep and Synaptic Homeostasis: Structural Evidence in Drosophila // Science. — 2011. — V. 332. — P. 1576—1581.
6.Matsuyoshi H., Kuniyasu H., Okumura M., et al. A 5-HT 4 receptor activation-induced neural plasticity enhaces in vivo reconstructs of enteric nerve circuit insult // Neurogastroenterol. 2010. — P. 806—813.
отправлен участнику