
Статья опубликована в рамках: XXXI Международной научно-практической конференции «Научное сообщество студентов XXI столетия. ЕСТЕСТВЕННЫЕ НАУКИ» (Россия, г. Новосибирск, 12 мая 2015 г.)
Наука: Медицина
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов

БИОХИМИЧЕСКИЕ ОСНОВЫ ПАТОГЕНЕЗА ШИЗОФРЕНИИ
Николаева Ольга Александровна
студент 2 курса лечебного факультета НГМУ, РФ, г. Новосибирск
E -mail: olg7252979@yandex.ru
Сычева Ирина Михайловна
научный руководитель, канд. хим. наук, доцент, кафедра медицинской химии НГМУ, РФ, г. Новосибирск
Первые описания шизофреноподобных симптомов встречались еще до нашей эры в папирусах Древнего Египта, и вот прошло более века с тех пор, как швейцарский психиатр Эйген Блейлер описал шизофрению как самостоятельное заболевание, но ее биологическая природа остается до сих пор загадкой. Богатство симптомов, вариантов течения делает это заболевание достаточно сложным для изучения, в то же время его лечение — процесс непростой и длительный. Несмотря на это, появление новых способов диагностики и исследования значительно облегчило поиск причины шизофрении, разработку новых методов терапии. Биохимические исследования показали, что в крови и биологических жидкостях у больных шизофренией изменен уровень важных для нормальной работы центральной нервной системы соединений, а также их метаболитов. В нашей работе будет представлено влияние нарушений обмена дофамина, серотонина и ГАМК, которое играет большую роль в патогенезе шизофрении.
I. Влияние дофамина
Дофамин относится к катехоламинам, он образуется под действием дигидроксифенилаланин — декарбоксилазы из дигидроксифенилаланина (ДОФА), который, в свою очередь, синтезируется из тирозина [2]. Рецепторы катехоламинов расположены, в основном, в плазматической мембране и являются метаботропными, зависимыми от G-белков. Примечательно, что реакция на гормон зависит от концентрации гормон-рецепторных комплексов, на которую влияет количество и гормона, и рецепторов. При длительном действии возникает снижение чувствительности рецепторов к гормону.
Сами рецепторы дофамина представлены двумя группами: D1 и D2-подобные [10], которые имеют некоторые отличия в механизмах действия. В основном, наибольшая концентрация рецепторов достигается в мезолимбических, кортикальных, нигростриальном путях.
Предполагается, что при шизофрении происходит избыточная активация мезолимбических и кортикальных путей. Это доказывается антипсихотическим действием нейролептиков (галоперидол, флюфеназин), которые, являясь антагонистами D2-подобных рецепторов, производят блокаду «позитивных» симптомов шизофрении — галлюцинаций, бреда, расстройства мышления. В то же время их действие довольно незначительно снижает «негативные» симптомы – плоскость аффекта, алогию и аволицию. Однако продолжительное действие данных нейролептиков может вызвать и побочное действие — паркинсонизм, в связи с длительной блокадой рецепторов в стриатуме [2]. Хотелось бы добавить, что сама смена «позитивной» и «негативной» симптоматики происходит на фоне изменений в дофаминовой системе: снижение ее активности ведет к снижению активности больного, депрессии, ослаблению когнитивных процессов, усилению негативной симптоматики, ее повышение же приводит к обратному эффекту.
II. Влияние гамма-аминомасляной кислоты
ГАМК является основным ингибиторным медиатором центральной нервной системы, и именно ей приписывают дефицит ингибирования, наблюдающийся при шизофрении [3]. Она широко распространена в коре головного мозга, стриатуме, таламусе, гиппокампе. ГАМКергические афферентные нейроны стриатума контролируют режим электрической активности дофаминовых нейронов в среднем мозге, при этом активация ГАМК-рецепторов приводит к подавлению выброса дофамина [8].
ГАМК синтезируется в организме из глутамата под действием ГАМК-декарбоксилазы, имеющей 2 изоформы — GAD65 и GAD67. Установлено, что именно GAD67 играет ключевую роль в регуляции синтеза ГАМК [4]. Затем ГАМК выбрасывается в синаптическую щель и связывается с постсинаптическими рецепторами, но часть ее обратно захватывается при помощи транспортера GAT-1 и инактивируется трансаминазой и дегидрогеназой [3].
Далеко не последнюю роль играет рилин — гликопротеин, который синтезируется интернейронами коры, гиппокампа и базальными ганглиями. После секреции он адгезируется на дендритах корковых пирамидных нейронов, окружая таким образом их «шипики» [9], которые играют важную роль в пластичности коры головного мозга. Рилин регулирует созревание дендритов, формирование их цитоскелета; он окружает тела и дендриты ГАМК-нейронов, способствуя тем самым высвобождению ГАМК [5].
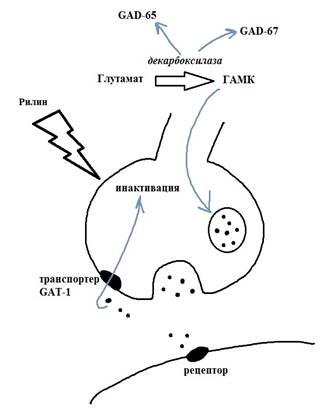
Рисунок 1. транспорт ГАМК в синапсе
Таким образом, все нарушения при шизофрении можно разделить на две группы: первая — касающиеся пресинаптических нейронов, их ферментов и транспортеров, вторая — касающиеся постсинаптических изменений.
Главным нарушением (и, пожалуй, наиболее распространенным среди всех изменений в транспорте ГАМК) первой группы является снижение экспрессии фермента GAD67 с белком рилином [7]. Снижение уровня мРНК можно обнаружить в 1, 2, 4 слоях коры, в таких зонах как префронтальная кора, передняя поясная и верхняя височная извилины, первичная моторная и зрительная области. Снижение экспрессии происходит также и вне коры – в гиппокампе, мозжечке и хвостатом ядре. Хотелось бы отметить, что данные нарушения не связаны со снижением самих ГАМКергических нейронов или их повреждением [6].
В ходе исследований выяснилось, что длительный прием агонистов дофамина приводит к еще большему угнетению GAD67, в то время как лечение нейролептиками повышает экспрессию этого фермента [8]. Это в очередной раз доказывает антагонизм ГАМКергической и дофаминергической систем.
К постсинамптическим нарушениям относят повышение экспрессии различных подтипов рецепторов, их субъединиц, которые имеют место быть во 2 и 3 слое префронтальной коры, гиппокампе, хвостатом ядре [6].
Таким образом, падение уровня ГАМК при различных нарушениях обуславливает развитие психозов с богатой продуктивной симптоматикой, тяжелые поведенческие и когнитивные изменения.
III. Влияние серотонина
Серотонин выполняет множество различных функций во многих тканях, что обуславливается наличием нескольких взаимодействующих между собой серотониновых подсистем. Так, например, подсистема, представленная нейронами головного мозга в ядрах шва, играет огромную роль в развитии всей нервной системы в целом в процессе онтогенеза, а во взрослом мозге — в сигнализации и настройке его многих отделов. Серотонин участвует в регуляции сексуального и пищевого поведения, агрессии, депрессии и тревожности.
Было проведено множество исследований по оценке количества меж- и внутриклеточного серотонина при шизофрении, однако результаты не совпадали: встречалось как повышение, так и понижение серотонина при данном заболевании. Позднее был обнаружен феномен полиморфизма генов, связанных с серотонинергической системой. Серотонин образуется из триптофана, а затем, упакованный в везикулы, транспортируется к синапсам. Из везикул он высвобождается в синаптическую щель, затем, попав в постсинаптический нейрон, он объединяется с рецепторами, а избыток его захватывается специальным белком и возвращается в пресинаптический нейрон. Под действием фермента моноаминооксидазы серотонин превращается в 5-гидроксииндолальдегид.
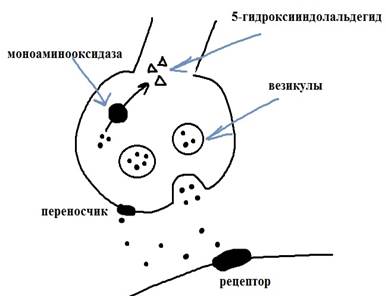
Рисунок 2. транспорт серотонина в синапсе
Наиболее изученными являются гены, кодирующие белок-переносчик и моноаминооксидазу. Участок гена-переносчика серотонина, прилегающий к промотору, (5-HTTLPR) содержит различное число повторов длиной 22 пары нуклеотидов каждая. У человека чаще встречаются аллели, имеющие либо 18 (длинный аллель), либо 16 повторов (короткий аллель). Если аллель короткий, то обратный захват серотонина снижался — внеклеточная концентрация росла, а внутриклеточная уменьшалась. Таким образом, у носителей короткого аллеля признаки тревожного ряда проявляются наиболее ярко.
В гене, кодирующем моноаминооксидазу, наиболее изучен полиморфизм в области промотора, обусловленный различным числом повторов длиной в 30 пар нуклеотидов. У человека существует пять видов аллелей с числом повторов от двух до пяти. Выяснилось, что некоторые аллели экспрессируются в два раза активнее. В результате в мозге во внеклеточном пространстве, уменьшается концентрация нейромедиатора, особенно на ранних стадиях онтогенеза, что приводит к нарушению формирования нормальных связей между нейронами.
Таким образом, описанный полиморфизм может влиять на клинические особенности течения данного заболевания. Например, симптомы депрессии свойственны носителям короткого аллеля 5-HTTLPR. Также имеются данные о более сильной эмоциональной реакции таких больных на собственные галлюцинаторные голоса, хотя в целом галлюцинации проявляются у пациентов с двумя длинными аллелями [1].
IV. Выводы
Таким образом, нами была рассмотрена биохимия патогенеза шизофрении, проанализировав данные, мы можем сделать следующие выводы:
1. В патогенезе данного заболевания огромную роль играют нарушения регуляторных механизмов с участием дофамина, серотонина и ГАМК
2. В разных участках нервной системы они вызывают различные эффекты
3. Данные три системы работают не раздельно, а в тесном контакте между собой
4. Многие нарушения биохимических механизмов обусловлены различными дефектами на генном уровне
Список литературы:
1.Голимбет В.Е., Алфимова М.В. Серотонин и шизофрения //Природа. — 2012. — № 8. — С. 34—38.
2.Кулинский В.И., Колесниченко Л.С. Катехоламины: биохимия, фармакология, физиология, клиника // Вопросы медицинской химии. — 2002. — т. 48, — № 1. — С. 44—67.
3.Нестерович А.Н., Голубович В.В., Гелда А.П. Дефицит ингибиторных систем головного мозга в рамках современных представлений о шизофрении // Военная медицина. — 2013. — № 2. — С. 123—131.
4.Asada H. Mice lacking the 65 kDa isoform of glutamic acid decarboxylase (GAD65) maintain normal levels of GAD67 and GABA in their brains but are susceptible to seizures / H. Asada [et al.] //Biochem. Biophys. Res. Commun. — 1996. — Vol. 229. — P. 891—895.
5.Grayson D.R. Reelin promoter hypermethylation in schizophrenia/ D.R. Grayson [et al.] // Proc. Natl Acad. Sci. USA. — 2005. — Vol. 102 (26). — P. 9341—9346.
6.Guidotti A. GABAergic dysfunction in schizophrenia: a new treatment on the horizon // Psychopharmacology (Berl.). — 2005. — Vol. 180. — P. 191—205.
7.Hashimoto T. Alterations in GABA-related transcriptome in the dorsolateral prefrontal cortex of subjects with schizophrenia / T. Hashimoto [et al.] // Mol. Psychiatry. — 2008. — Vol. 13. — P. 147—161.
8.Kalkman H.O. GAD67: the link between the GABA-deficit hypothesis and the dopami-nergic- and glutamatergic theories of psychosis / H.O. Kalkman, E. Loetscher // J. Neural. Transm. — 2003. — Vol. 110. — P. 803—812.
9.Noh J.S. DNA methyltransferase 1 regulates reelin mRNA expression in mouse primary cortical cultures / J.S. Noh [et al.] //Proc. Natl. Acad. Sci. USA. — 2005. — Vol. 102 (5). — P. 1749—1754.
10.Strange Pb.G. Dophamine receptors // Tocris Rev. — 2000. — № 78. — p. 1—6.
дипломов