
Статья опубликована в рамках: XXIV Международной научно-практической конференции «Научное сообщество студентов XXI столетия. ЕСТЕСТВЕННЫЕ НАУКИ» (Россия, г. Новосибирск, 28 октября 2014 г.)
Наука: Биология
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов

ДИНАМИКА НАРАСТАНИЯ ОДНОЛЕТНИХ ПОБЕГОВ ИВЫ ШЕРСТИСТОПОБЕГОВОЙ (SALIX DASYCLADOS WIMM.) В УСЛОВИЯХ БРЯНСКОГО ЛЕСНОГО МАССИВА
Шилина Елена Александровна
студент 5 курса, кафедра биологии БГУ, РФ, г. Брянск
E -mail: dasyclados@mail.ru
Афонин Алексей Алексеевич
научный руководитель, д-р с.-х. наук, профессор БГУ, РФ, г. Брянск
Ива шерстистопобеговая (Salix dasyclados Wimm. 1849 = Salix gmelinii Pall. 1789) — широко известный представитель секции Vimen Dum. (Viminella Ser.) подрода Vetrix Dum. рода Salix L. [2; 8; 9].
Биологический потенциал ивы шерстистопобеговой — способность к аутовегетативному размножению зимними черенками и высокая продуктивность — используется при создании противоэрозионных, биоремедиационных, декоративных насаждений, а также разнообразных культур плантационного типа. В последние десятилетия ива шерстистопобеговая рассматривается как перспективная энергетическая культура [2; 3; 4; 5; 6].
Ареал ивы шерстистопобеговой — евроазиатского бореального типа; в европейской части России южная граница ареала проходит в Верхне-Днепровском районе, а на территории Брянского лесного массива (БЛМ) — примерно по 53 параллели [8]. Эти краевые популяции ивы шерстистопобеговой входят в состав ассоциаций Salicetum triandro-viminalis (Tx. 1931) Lohm. 1952 (в понижениях центральной и прирусловой поймы), Salicetum albae Issl. (1924) 1926 (прирусловые ивняки) и Salicetum cinereae Pass. 1956 (на сырых внепойменных лугах и лесных полянах) [2]. В указанных эдафо-фитоценотических условиях высокопродуктивные клоны ивы шерстистопобеговой могут использоваться для создания полифункциональных насаждений, в которых наиболее полно реализуется эколого-генетический потенциал вида [3; 4; 5; 6].
В благоприятных почвенно-климатических условиях фактическая продуктивность плантационных культур ивы шерстистопобеговой составляет 10,6 т/га×год (около 150 ГДж/га×год), что примерно в 1,5 выше продуктивности других энергетических ив [7]. Известно [1; 10], что высокий и стабильный урожай биомассы ив может быть обеспечен путем создания поликлональных насаждений, образованных биотипами, различающимися по ритмам нарастания побегов. В то же время, закономерности развития однолетних побеговых систем ивы шерстистопобеговой изучены недостаточно [3; 4; 5; 6], что и обусловливает актуальность данного исследования.
Цель исследования : Выявить закономерности сезонной динамики нарастания однолетних побегов ивы шерстистопобеговой.
Задачи исследования :
1. Выявить периодичность нарастания побегов ивы шерстистопобеговой на одновозрастном, генетически выровненном материале, культивируемом в однородных условиях.
2. Изучить общие закономерности сезонной динамики нарастания сильных побегов высокопродуктивных сеянцев ивы шерстистопобеговой.
3. Выявить разнообразие ритмов сезонной динамики нарастания побегов у разных биотипов ивы шерстистопобеговой.
4. Разработать рекомендации по рациональному использованию существующего разнообразия ритмов сезонной динамики нарастания побегов для создания устойчивых высокопродуктивных насаждений.
Материал и методы исследования
В качестве материала для данного исследования использовалась семья ивы шерстистопобеговой da 09-х, полученная в культуре [3]. Исходная семья da 01-х была получена путем посева семян, собранных в природе 20.05.2001 с пестичной особи Salix dasyclados в центральной пойме р. Десны (на территории Снежетьско-Деснянского низменного (долинно-речного) ландшафта БЛМ), в ассоциации Salicetum triandro-viminalis (Tx. 1931) Lohm. 1952 в точке с координатами 34,371874º в.д., 53,220569º с.ш. Полученные сеянцы культивировались в салицетуме Брянского государственного университета (ТЛУ D3). Весной 2009 г. одна из рамет высокопродуктивного пестичного клона da 01-7 была опылена пыльцой высокопродуктивного тычиночного клона ивы корзиночной (S. viminalis). В результате было получено небольшое количество семян, из которых были выращены сеянцы семьи da 09-х. Все сеянцы обладали типичными чертами S. dasyclados, что позволяет допустить их апомиктическое происхождение. Полученные сеянцы da 09-х также культивировались в салицетуме Брянского государственного университета. Расположение — по тригонально-центрированной схеме с расстоянием между посадочными местами 2 м.
Для данного исследования использовались сеянцы da 09-2, da 09-5 (в дальнейшем da 2, da 5) с высоким годичным приростом двух первых замещающих побегов (110…185 см). Каждая такая пара побегов с каждой особи рассматривалась как побеговая система. Материал собирался на протяжении вегетационного сезона 2012 г. За дату начала вегетации было принято 11 мая (начало интенсивного роста побегов), за дату окончания — 22 августа (к этому времени развитие побегов практически завершается). Каждые 4…7 суток на каждом сеянце замеряли длину (h, см) двух первых замещающих побегов. Обработка полученных результатов производилась с помощью редактора электронных таблиц Microsoft Excel (MS Office 2003, 2010).
Текущий среднесуточный линейный прирост побегов (Δh(t), см/сут) для каждой расчетной даты (t = (tk+1+tk–1) / 2) рассчитывался по формуле:
Δh(t)=(hk+1–hk–1)/(tk+1–tk–1),
где: h — текущая длина лидерного побега (см),
k — порядковый номер наблюдения,
t — порядковый день вегетационного сезона. В дальнейшем среднесуточный прирост Δh(t) для каждой расчетной даты t обозначался в сокращенной форме — Δh.
Для выявления различий в ритмах нарастания побегов использовался визуальный анализ рядов динамики Δh=f(t).
Результаты и обсуждение
Сравнительная динамика линейного прироста побеговых систем сеянцев da 2 и da 5показана на рисунке № 1.
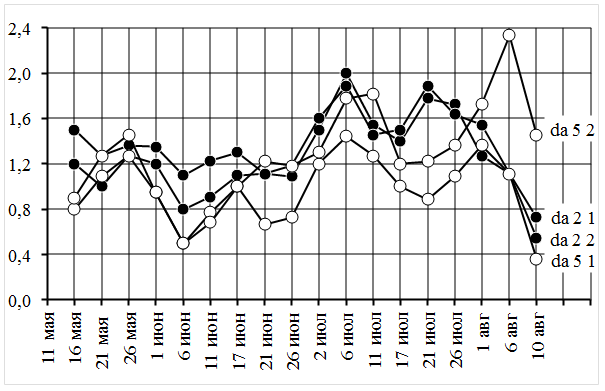
Рисунок 1. Сравнительная динамика линейного прироста побеговых систем
По оси абсцисс расчетные даты 2012 г., по оси ординат среднесуточный прирост (Δh, см/сут). Справа указаны сеянцы и номера побегов.
Сравнение рядов сезонной динамики среднесуточных приростов длины побегов (Δ h) показало, что нарастание побегов на протяжении всего периода вегетации происходит неравномерно.
В первой половине вегетационного периода (до середины июля) развитие побегов на обоих сеянцах протекало сходным образом. Весенний локальный максимум среднесуточного прироста (Δ h = 0,8…1,5 см/сут) зафиксирован 16…26 мая. После прохождения этого максимума среднесуточный прирост снизился до абсолютного минимума (Δ h = 0,5…1,1 см/сут) 6 июня. Далее, к 17 июня Δ h увеличился (до 1,0…1,3 см/сут), а затем несколько снизился (Δ h = 0,7…1,2 см/сут). После 26 июня Δ h резко увеличился и достиг общего среднелетнего максимума 6 июля (Δ h = 1,4…2,0 см/сут). После прохождения этого максимума среднесуточный прирост снизился к 17 июля до среднелетнего локального минимума (Δ h = 1,0…1,5 см/сут). С этого момента динамика развития побегов приобрела индивидуальный характер.
Среднесуточный прирост побегов сеянца da 5 стабилизировался на уровне Δ h = 0,9…1,4 мм/сут до 26 июля, а затем вырос до Δ h = 1,4…2,3 мм/сут в первой пятидневке августа (позднелетний максимум). После прохождения этого максимума среднесуточный прирост побегов сеянца da 5 резко снизился до Δ h = 0,4…1,5 см/сут к 10 августа, и развитие побегов завершилось к 22 августа 2012 г. Побеги сеянца da 2, напротив, активизировали свой рост после прохождения среднелетнего локального минимума, и к 21 июля среднесуточный прирост побегов достиг 1,8…1,9 мм/сут (позднелетний максимум). После прохождения этого максимума среднесуточный прирост побегов сеянца da 2 постепенно снижался до Δ h = 0,5…0,8 мм/сут к 10 августа, и развитие побегов прекратилось к 17 августа 2012 г.
Таким образом, на одновозрастном, генетически выровненном материале установлено, что побеговые системы ивы шерстистопобеговой характеризуются наличием трех локальных максимумов среднесуточного линейного прироста, причем третьи максимумы второй половины вегетационного периода несколько смещены относительно друг друга. На этом основании можно предположить, что выявленные биоритмы представляют собой комбинации гармонических колебаний с разными периодами (длинно- и короткопериодические) и с различной амплитудой. Тогда каждый побег может рассматриваться как осциллятор — колебательная система, в которой изменчивость ритмов прироста побегов определяется в большей степени эндогенными факторами, т. е. генетически обусловленными программами морфогенеза [2].
Выводы и рекомендации
На одновозрастном, генетически выровненном материале, культивируемом в однородных условиях на протяжении вегетационного периода 2012 г., установлено следующее.
1. Сезонная динамика нарастания побегов ивы шерстистопобеговой характеризуется закономерным чередованием периодов интенсивного и ослабленного роста.
2. Сезонная динамика нарастания побегов ивы шерстистопобеговой отличается высоким среднесуточным приростом в июле — начале августа. Таким образом, высокопродуктивные сеянцы ивы шерстистопобеговой максимально используют ресурсы второй половины лета.
3. На протяжении вегетационного периода выявлено четыре периода максимального прироста, разделенных периодами относительного покоя.
4. В первой половине вегетационного периода влияние индивидуальных особенностей сеянцев на динамику нарастания побегов не установлено. Во второй половине этого же вегетационного периода выявлено смещение позднелетних максимумов среднесуточного прироста, обусловленное индивидуальными особенностями сеянцев.
5. Разнообразие сезонных ритмов нарастания однолетних побегов ивы шерстистопобеговой рекомендуется учитывать при проектировании устойчивых высокопродуктивных насаждений комплексного назначения.
Список литературы:
1.Афонин А.А. Методологические принципы создания устойчивых высокопродуктивных насаждений ив (на примере автохтонных видов Salix Брянского лесного массива). Брянск: РИО Брянского гос. ун-та, 2005. — 146 с.
2.Афонин А.А. Сравнительная морфодинамика однолетних побегов ив Брянского лесного массива. Брянск: Изд-во Курсив, 2011. — 145 с.
3.Афонин А.А. Динамика нарастания лидерных побегов сеянцев ивы шерстистопобеговой (Salix dasyclados Wimm.) // Лісове і садово-паркове господарство. К.: ЦП Компринт, — 2012. — № 2. — C. 5—24.
4.Афонин А.А. Формовое разнообразие ивы шерстистопобеговой (Salix dasyclados Wimm. = Salix gmelinii Pall.) на территории Восточной Европы // Вестник БГУ. Естественные и точные науки. — 2013. — № 4. — С. 57—60.
5.Афонин А.А., Анищенко Л.Н. Разнообразие генетических программ морфогенеза в популяциях Salix dasyclados Wimm. (Salix gmelinii Pall.) // Ученые записки института сельского хозяйства и природных ресурсов НовГУ. Т. 18. Вып. 2. Великий Новгород: НовГУ, 2010. — С. 63—67.
6.Афонин А.А., Борздыко Е.В., Анищенко Л.Н. Эколого-генетический потенциал природных популяций ивы шерстистопобеговой // Фундаментальные науки и практика: Сб. научн. тр. 3-й Междунар. телеконф. Томск: СибГМУ, 2010. — С. 159—160.
7.Логинова Л.А. Продуктивность и энергетический потенциал ивовых ценозов на примере Воронежской области. Автореф. дис… канд. биол. наук. Воронеж, 2010. — 19 с.
8.Скворцов А.К. Ивы СССР. М.: Наука, 1968. — 262 с.
9.Belyaeva I., Sennikov A. Tipification of Pallas′ names in Salix // Kew Bulletin. — 2008. — Vol. 63. — P. 277—287.
10.McCracken A.R. Interaction of willow (Salix) clones growing in mixtures // Ann. Appl. Biol. — 1998. — 132 Suppl. — P. 54—55.
дипломов