
Статья опубликована в рамках: XVI-XVII Международной научно-практической конференции «Наука вчера, сегодня, завтра» (Россия, г. Новосибирск, 06 октября 2014 г.)
Наука: Биология
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов

Статья опубликована в рамках:
Выходные данные сборника:
ПОТЕНЦИАЛЬНО ТОКСИЧНЫЕ ДИАТОМЕИ КОМПЛЕКСА PSEUDO -NITZSCHIA PSEUDODELICATISSIMA/СUSPIDATA ИЗ ЗАЛИВА ПЕТРА ВЕЛИКОГО ЯПОНСКОГО МОРЯ
Стоник Инна Валентиновна
канд. биол. наук, старший научный сотрудник Институтa биологии моря имени А.В. Жирмунского ДВО РАН, РФ, г. Владивосток
E -mail: stonikiv@mail.ru
Работа выполнена при финансовой поддержке ДВО РАН (проект № 12- III-A-06-096)
Пищевое отравление, вызванное токсичными диатомеями, получило название Amnesic Shellfish Poisoning (ASP) — амнезическое отравление моллюсками [5, с. 978]. Этот тип отравления связан с накоплением нейротоксина (домоевой кислоты), продуцируемой диатомовыми водорослями, в двустворчатых моллюсках и некоторых других морских беспозвоночных и последующей передачей токсина по пищевым цепям, что может привести к гибели морских млекопитающих, птиц и человека. В настоящее время 14 видов Pseudo-nitzschia известны как потенциальные продуценты домоевой кислоты [6, с. 172]. Для большинства таких видов показано существование как токсичных, так и нетоксичных клонов (клеточных линий). Среди видов — возбудителей ASP в северо-западной части Японского моря наиболее часто токсичные «цветения» вызывают представители сложных видовых комплексов P. pseudodelicatissima/cuspidata, P. delicatissima и P. Pungens [9, с. 132]. Представители первого из указанных комплексов достигают наиболее высокой плотности в заливе Петра Великого Японского моря. Однако морфология и таксономия представителей данного комплекса из российских вод Японского моря изучены недостаточно.
Цель данной работы — изучить особенности морфологии, таксономический состав и плотность представителей комплекса P. pseudodelicatissima/cuspidata из залива Петра Великого Японского моря.
Материалом послужили батометрические пробы фитопланктона, собранные в Амурском и Уссурийском заливах (вторичные заливы зал. Петра Великого) Японского моря в разные сезоны за период с 2008 г. по 2013 г. Круглогодичное изучение фитопланктона проводили на монитринговой станции в районе м. Красного в Амурском заливе. Батометрические пробы объемом 1 л отбирали с поверхности воды на разных горизонтах до глубины 30 м. Один литр пробы фиксировали раствором Утермеля и концентрировали методом осаждения [1, с. 84] и/или обратной фильтрации через нуклеопоровые фильтры с диаметром пор 2 мкм. Материал фиксировали раствором Утермеля. Плотность клеток подсчитывали в счетной камере (Sedgewick-Rafter) объемом 1 мл. Под плотностью фитопланктона понимали число клеток в литре воды. Изучение материала проводили с помощью светового микроскопа Olympus BX-41 в течение 2—3 месяцев после отбора проб, а также трансмиссионного электронного микроскопа (ТЭМ, JEM-100 S). При подготовке материала к ТЭМ использовали традиционные методики очистки панцирей, включающие воздействие кислот и центрифугирование с дистиллированной водой. Для изучения материала с помощью ТЭМ каплю отмытой пробы наносили на бленды с формваром и высушивали на воздухе. В работе использована стандартная терминология, принятая для диатомовых водорослей [4, с. 344; 10, с. 152].
Комплекс P. pseudodelicatissima/сuspidata характеризуется высоким таксономическим разнообразием и объединяет виды, имеющие относительно узкие (ширина 1,5—3 мкм) створки линейной или ланцетной формы, центральный узелок и штрихи со структурой из одного ряда ареол (пороидов) [7, с. 797]. В результате недавних исследований в пределах этого комплекса на основе морфологических, молекулярно-генетических и репродуктивных данных были описаны новые псевдокриптические виды [8, с. 436]. Известно, что внутри этой группы видов, идентификация сложна и основана на комбинации нескольких морфологических признаков (плотность фибул, штрихов створки и первого пояскового ободка, а также число секторов пороидов створки) [8, с. 450]. В результате проведенных нами исследований в заливе Петра Великого обнаружены следующие представители указанного комплекса: P. caciantha, P. calliantha, P. cuspidata, Pseudo-nitzschia sp. Их морфометрические характеристики приведены в таблице 1.
Морфометрические характеристики изученных нами экземпляров видов P. caciantha и P. cuspidata согласуются с соответствующими данными, имеющимися в научной литературе (таблица 1). Изученные нами клетки P. calliantha характеризовались наличием основных диагностических признаков вида — разделение гимена пороидов на 7—10 секторов и центральную перфорированную область, а также структура штрихов вальвокопулы из 2—3 первальварных рядов [8, с. 436]. Однако необходимо отметить небольшие отклонения значений некоторых морфологических признаков у наших экземпляров P. calliantha от приведенных в литературе. Так, средняя плотность фибул и штрихов створки у наших экземпляров оказались выше соответствующих значений, приведенных для других клонов этого вида (таблица 1). Высокая морфологическая изменчивость P. calliantha характерна для представителей вида из разных местообитаний и может объясняться его значительной генетической изменчивостью [7, с. 811]. Морфология изученных нами клеток Pseudo-nitzschia sp. (таблица 1) не позволила отнести его к какому-либо из известных видов комплекса Pseudo-nitzschia pseudodelicatissima/cuspidate [2, с. 198; 3, с. 491; 7, с. 800; 8, с. 436]. Этот морфотип по числу секторов пороидов (2—7) близок к видам P. mannii (2—7) и P. hasleana (2—6). Однако плотность штрихов створки (43—49 в 10 мкм) и штрихов вальвокопулы (54—65 в 10 мкм) оказались существенно выше таковых для других видов комплекса. Для идентификации этого таксона необходимы дальнейшие исследования с привлечением морфологических и молекулярно-генетических данных.
Таблица 1.
Морфометрические характеристики представителей комплекса P . pseudodelicatissima/сuspidata, найденных в заливе Петра Великого, и данные, имеющиеся в научной литературе
|
Вид |
Ширина створки ( мкм) |
Плот-ность фибул в 10 мкм |
Плот-ность штрихов створки в 10 мкм |
Число секторов пороидов на створке |
Плот-ность штрихов пояска в 10 мкм |
|
P . caciantha (наши измерения) |
2,6-3,2 2,9 ± 0,2 |
14-19 16,5 ± 1,8 |
30-32 31,3 ± 0,5 |
2—5 |
36-38 35,4 ± 1,6 |
|
P . caciantha [7, с. 800] |
2,2—3 |
18—23 |
33—37 |
2—6 |
35—38 |
|
P . calliantha (наши измерения) |
1,4−2 1,6 ± 0,2 |
25−27 25,7 ± 1 |
39−45 42,2 ± 2,5 |
7—10 |
42−43 42,3± 0,6 |
|
P. calliantha [8, с. 438] |
1,4-1,8 |
15-22 |
34-39 |
7—10 |
42-48 |
|
P. cuspidata (наши измерения) |
1,4−2 1,7 ± 0,2 |
19−24 21,3 ± 2,4 |
35−43 38,8 ± 3,3 |
2 |
48 |
|
P . cuspidata [8, с. 438 ] |
1,4—2 |
19—25 |
35—44 |
2 |
47—53
|
|
Pseudo -nitzschia sp. (наши измерения) |
1,5-2,2 1,7± 0,2 |
23-29 24± 3,8 |
43-49 47± 1,9 |
2—7 |
54-65 58,3± 6,2 |
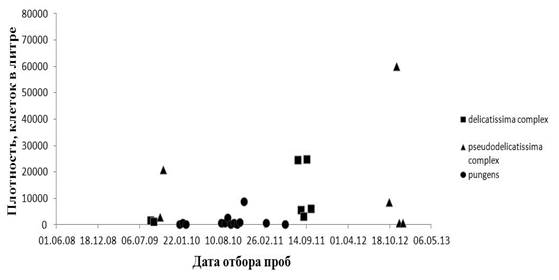
Рисунок 1. Динамика плотности представителей комплексов P . pseudodelicatissima/cuspidata, P. delicatissima и P. pungens на мониторинговой станции в Амурском заливе в период с июня 2008 г. по май 2013 г.
Анализ многолетней динамики плотности видов рода Pseudo-nitzschia в Амурском заливе показал, что в исследуемый период представители комплекса Pseudo-nitzschia pseudodelicatissima/cuspidatа достигали наиболее высокой плотности по сравнению с видами комплекса Pseudo-nitzschia delicatissima и P. pungens (рисунок 1). Так, в период с 2009 г. по 2012 г. плотность представителей комплекса Pseudo-nitzschia pseudodelicatissima/cuspidatа изменялась в пределах от 25 до 60 тыс. кл/л. Массовое развитие видов комплекса определяло осеннее цветение потенциально токсичных диатомей в октябре-ноябре 2012 г.
Таким образом, в результате изучения таксономического состава представителей комплекса Pseudo-nitzschia pseudodelicatissima/cuspidata из залива Петра Великого идентифицированы следующие таксоны: P. cf. caciantha, P. calliantha, P. cuspidata, Pseudo-nitzschia sp. Вид P. cuspidatа впервые найден в дальневосточных морях России. Установлена высокая плотность видов комплекса (до 60 тыс. кл/л) в фитопланктоне Амурского залива Японского моря.
C писок литературы:
1.Федоров В.Д. О методах изучения фитопланктона и его активности. М.: Изд-во МГУ, 1979 — 168 с.
2.Amato A., Kooistra W.H.C.F., Levialdi J.H., Mann D.G., Proshold T., Montresor M. Reproductive isolation among sympatric cryptic species in marine diatoms // Protist. — 2007. — Vol. 158. — P. 193—207.
3.Amato A., Montresor, M. Morphology, phylogeny, and sexual cycle of Pseudo-nitzschia mannii sp. nov. (Bacillariophyceae): a pseudo-cryptic species within th P. pseudodelicatissima complex // Phycologia — 2008. — Vol. 47. — P. 487—497.
4.Anonymous. Proposals for a standardization of diatom terminology and diagnoses // Nova Hedwigia, Beihefte. — 1975. — Vol. 53. — P. 323—354.
5.Bates S.S. Domoic-acid-producing diatoms: another genus added! // J. Phycol. — 2000. — Vol. 36. — P. 978—983.
6.Lelong A., Hegaret H., Soudant P., Bates S.S. Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amnesic shellfish poisoning: revisiting previous paradigms // Phycologia. — 2012. — Vol. 51, — № 2. — P. 168—216.
7.Lundholm N., Moestrup Ø., Hasle G.R., Hoef-Emden K.A. A study of the Pseudo-nitzschia pseudodelicatissima/P. cuspidata complex (Bacillariophyceae): what is P. pseudodelicatissima? // Journal of Phycolology. — 2003. — Vol. 39. — P. 797—813.
8.Lundholm N., Bates S.S., Baugh K.A., Bill B.D., Connell L.B., Leger C., Trainer V.L. Cryptic and pseudo-cryptic diversity in diatoms — with descriptions of Pseudo-nitzschia hasleana sp. nov. and P. fryxelliana sp. nov. // Journal of Phycolology. — 2012. — Vol. 48. — P. 436—454.
9.Round F.E., Crawford R.M., Mann D.G. The Diatoms. Biology and morphology of genera. Cambridge: Cambridge University Press, 1990. — 747 pp.
10.Stonik I.V., Orlova T.Yu., Lundholm N. Diversity of Pseudo-nitzschia H. Peragallo from the western North Pacific // Diatom Research. — 2011. — Vol. 26. — P. 121—134.
дипломов