
Статья опубликована в рамках: IX Международной научно-практической конференции «Наука вчера, сегодня, завтра» (Россия, г. Новосибирск, 10 февраля 2014 г.)
Наука: Сельскохозяйственные науки
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов

О ПРОЯВЛЕНИИ МЕЖВИТАМИННЫХ ВЗАИМОДЕЙСТВИЙ РЕТИНОЛА И АЛЬФА-ТОКОФЕРОЛА В ТКАНЕВОМ ДЫХАНИИ ПЕЧЕНИ КУР
Костюк Инна Александровна
канд. с.-х. наук, и. о. доцента кафедры нормальной и патологической физиологии Харьковской государственной зооветеринарной академии, Украина, Харьковcкая обл.,пгт Малая Даниловка
Е-mail:
В связи с известными межвитаминными взаимодействиями ретинола и альфа-токоферола, а также дополнительным применением в питании людей и в рационах животных [1; 3; 8; 9] целесообразно изучение их общего влияния на энергетический аппарат клетки. В работе изучались межвитаминные взаимодействия мембранотропных веществ ретинола (витамин А) и альфа-токоферола (витамин Е) в процессах тканевого дыхания в печени кур.
Цель работы — определить особенности процессов тканевого дыхания и окислительного фосфорилирования в митохондриях печени при совместном применении в рационе повышенных доз витаминов А и Е.
В рацион кур породы Род-Айленд в возрасте 180 дней вводили дополнительно витамины А и Е в виде масляных препаратов, в повышенных дозах и различных сочетаниях (табл. 1). В рацион кур контрольной группы вводили добавки в рекомендуемых дозах [4; 6]. После 40 дней применения добавок витаминов А и Е, во время наибольшего их накопления в яичном желтке и печени, определяли показатели дыхательной активности митохондрий печени: скорости дыхания в разных метаболических состояниях митохондрий (V2, V3, V4, Vднф), коэффициент дыхательного контроля (ДК), интенсивность и эффективность фосфорилирования (ИФ и АДФ/О). После закладки яиц для инкубации и получения цыплят от экспериментального поголовья, определяли такие же показатели у цыплят суточного возраста.
Методы исследования: выделение митохондрий из клеток печени методом дифференциального центрифугирования и регистрация поглощения кислорода митохондриями в инкубационной среде полярографическим методом [5, с. 92], определение витаминов А и Е с применением фотоколориметрических и хроматографических методов [5].
Результаты.
Эффекты витаминов А и Е в процессах тканевого дыхания имели отличия у птицы разного возраста. Негативное влияние накопления витаминов А и Е в печени на энергетические процессы в митохондриях было выявлено в нашем опыте только у суточных цыплят и у кур в возрасте 330 дней. А вот у кур возрастом 220 дней, которые потребляли эти витамины в высоких дозах на протяжении 40 дней, установлена стимуляция окисления субстратов митохондриями и активизация процессов окислительного фосфорилирования.
Межвитаминные взаимодействия в процессах накопления витаминов А и Е у кур после 40 дней потребления с кормом, проявлялись в следующем: увеличение концентрации витамина А в печени кур в 3 и 4 группах сопровождалось снижением концентрации витамина Е в 2—3 раза. Накопление витамина Е в печени способствовало росту концентрации витамина А (табл. 1).
Таблица 1.
Накопление витаминов А и Е в печени кур в зависимости от их поступления в организм с кормом (возраст 220 дней, M±m, n=5)
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Примечание: * — p<0,05; ** — p<0,01 — разница достоверна в сравнении с показателями 1 группы
Наибольшее содержание витамина А в печени определено у кур 4 группы, которые получали его в 40-кратной дозе (400 тыс. МЕ/кг корма) и добавку витамина Е (10 мг/кг корма). Одновременно при этом происходило снижение концентрации витамина Е в печени в 3,5 раза. У кур 7 группы, которые получали 20-кратную дозу витамина Е (200 мг/кг корма) и рекомендованную дозу витамина А (10 тыс. МЕ/кг), концентрация альфа-токоферола в печени повысилась почти в 10 раз.
Накопление витаминов А и Е в печени кур повлияло на дыхательную активность митохондрий. Снижение скорости окисления 2-оксоглутарата V2 на 46,7 % (p<0,05) отмечено в 4-й группе кур (табл. 2) при потреблении ими витамина А в количестве 400 тыс. МЕ/кг и снижении концентрации витамина Е, а также у кур 5 группы, которые не получали добавку витамина Е с кормом.
Наивысшая скорость V2 при окислении митохондриями сукцината была определена в 6 и 7 группах, где доза витамина Е составляла 100 и 200 мг/кг, соответственно, а доза витамина А — 10 тыс. МЕ/кг. Так, у кур 7 группы интенсивность окисления сукцината повысилась на 115,4 %, а в 6 группе этот показатель повысился на 78,9 % (р<0,05). У кур 5 группы, которые не получали добавку витамина Е, скорость V2 была ниже, чем в 6 и 7 группах.
Таблица 2.
Показатели дыхательной активности митохондрий печени кур в связи с потреблением витаминов А и Е с кормом (возраст 220 дней, M±m, n=5)
|
|
||||||
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Примечание: * — p<0,05; ** — p<0,01 — разница достоверна в сравнении с показателями 1 группы
Сравнивая скорости окисления 2-оксоглутарата Vднф (в присутствии разобщителя реакций дыхания и фосфорилирования динитрофенола - 2,4-ДНФ) следует отметить, что высокая скорость была установлена в 6 и 7 группах, где дозы витамина Е составляли 100 и 200 мг/кг. Так, у кур 7 группы Vднф при окислении 2-оксоглутарата в 2,9 раз превышала контроль. При окислении сукцината максимальная скорость Vднф в данном опыте установлена у кур 6 группы. Это отображает активизацию свободного окисления субстратов, как 2-оксоглутарата, так и сукцината, с увеличением концентрации витамина Е в печени кур в 7—10 раз.
Под действием витамина А также происходило увеличение скорости Vднф. Так, у кур 3 группы скорость Vднф при окислении 2-оксоглутарата повысилась на 60,5 % (р<0,05). Следовательно, накопление в течение 40 дней витамина А в печени кур, как и витамина Е, способствовало активизации свободного окисления субстратов. Такие изменения скорости окисления субстратов в разобщённом состоянии происходили при общем росте дыхательной активности митохондрий и значительном накопления этих витаминов в печени.
Следует заметить, что в отсутствие добавки витамина А (2 группа) и применении витамина Е (10 мг/кг) на протяжении 40 дней, концентрация витаминов А и Е приближалась к показателям контрольной группы, что можно объяснить либо эффективным использованием каротинов корма, либо перераспределением витамина А между органами птицы и более активным депонированием в печени под действием витамина Е. Другая ситуация складывается в отсутствие добавки витамина Е и применении гарантирующей добавки витамина А — 10 тыс. МЕ/кг (5 группа): значительно снижается дыхательный контроль при окислении сукцината и активизируется свободное окисление 2-оксоглутарата, растут скорости V4 и Vднф. Это свидетельствует о потребности в витамине Е для эффективного сопряжения процессов дыхания и фосфорилирования.
Дыхательный контроль (ДК) по Ларди, соотношение скоростей V3 и V2, характеризует потенциальную активность дыхательной цепи в условиях наивысшего сопряжения дыхания и окислительного фосфорилирования. При окислении 2-оксоглутарата ДК по Ларди повышался как под действием витамина А в 4 группе (от 2,82 ед. в контроле до 5,55 ед.), так и под действием витамина Е в 7 группе (от 2,82 ед. до 4,36 ед.).
Если сравнить интенсивность фосфорилирования (ИФ) 2 группы кур и показатели 3 и 4 групп, которые получали 100 и 400 тыс. МЕ/кг витамина А, то можно отметить, что ИФ в этих группах ниже, чем в 2 группе (рис. 1). Такие изменения могут быть связаны со значительным уменьшением концентрации витамина Е в печени. Это предположение подтверждается изменениями ИФ у кур 5 и 6 групп. Так, при окислении 2-оксоглутарата митохондриями печени, ИФ у кур 6 группы (доза витамина Е 100 мг/кг) значительно превышает этот показатель в 5 группе кур, которые не получали витамин Е.
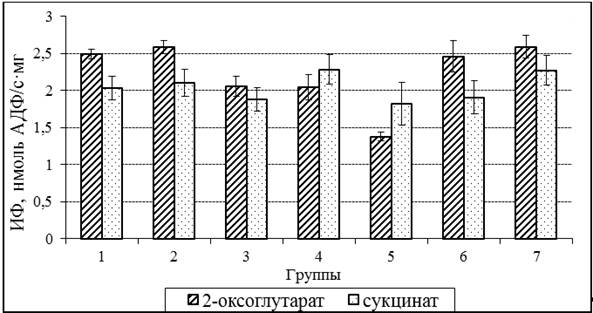
Рисунок 1. Интенсивность фосфорилирования в митохондриях печени кур при различном содержании А и Е в рационе
Эффективность фосфорилирования (АДФ/О) при окислении 2-оксоглутарата митохондриями снижалась, на 40,4 % (р<0,05), в 5 группе кур, которые не получали дополнительно витамин Е (рис. 2). Отмечено позитивное влияние витаминов А и Е на АДФ/О при окислении обоих субстратов в 3 и 6 группах.
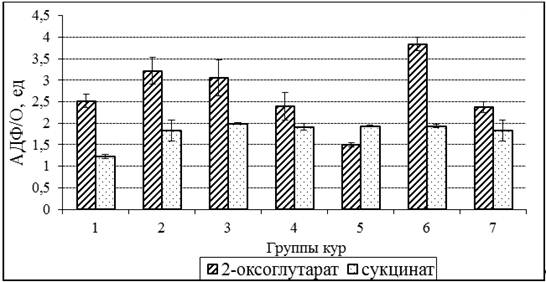
Рисунок 2. Эффективность фосфорилирования в митохондриях печени кур при различном содержании витаминов А и Е в рационе
Пик концентрации витаминов А и Е в яичном желтке у кур, которые получали добавки этих витаминов установлен на 35-й день опыта (рис. 3, рис. 4). Под действием высоких доз витамина А снижалось накопление витамина Е в яичном желтке. Однако, при повышении содержания витамина Е в желтке — концентрация витамина А также возростала.
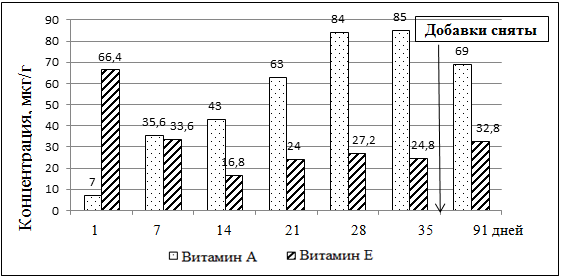
Рисунок 3. Динамика накопления витаминов А и Е в яичном желтке от 4 группы кур (доза витамина А — 400 тыс. МЕ/кг)
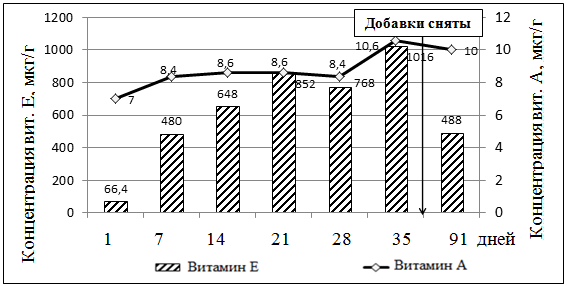
Рисунок 4. Динамика накопления витаминов А и Е в яичном желтке от 7 группы кур (доза витамина Е — 200 мг/кг)
Применение витамина А в рационе кур-несушек в количестве 100 тыс. МЕ/кг корма способствовало активизации окисления 2-оксоглутарата митохондриями печени цыплят суточного возраста, но при этом у цыплят снижались скорости окисления сукцината, а также уменьшался ДК. При применении витамина А в количестве 400 тыс. МЕ/кг корма замедлялось окисление 2-оксоглутарата и сукцината в митохондриях печени суточных цыплят, снижались АДФ/О и ИФ.
В результате применения в рационе кур-несушек витамина Е в 20-кратной дозе (200 мг/кг корма) он оказывал неблагоприятное влияние на тканевое дыхание в печени цыплят суточного возраста, поскольку активизировались процессы свободного окисления сукцината, замедлилось использование митохондриями 2-оксоглутарата, происходило разобщение процессов дыхания и фосфорилирования.
Обсуждение результатов.
Известно, что ретиноевая кислота найдена в составе митохондриального гена GRIM-19, определенного как фактор апоптоза, который одновременно кодирует последовательность субъединиц I комплекса ДЦ-NADH: убихинон-оксидоредуктазы [7]. Следовательно, активация окисления НАД-зависимых субстратов, в частности 2-оксоглутарата, под действием ретиноевой кислоты может привести к избыточному образованию сукцината и стимуляции его свободного окисления. Следовательно, повышение дыхательной активности митохондрий печени кур, установленное в наших опытах, может быть индуцировано двумя путями: 1) в результате влияния витамина А на I участок ДЦ; 2) его воздействием на активность ПОЛ и структуру мембран.
Существует предположение [2], что витамин Е влияет на процессы окислительного фосфорилирования, связанные с I участком ДЦ, известно о его взаимодействии с убихиноном Q. Однако прямых окислительно-восстановительных взаимодействий между НАД и альфа-токоферолом до настоящего времени не установлено. Результаты наших исследований, также, отображают влияние витамина Е на окисление НАД-зависимого субстрата 2-оксоглутарата и, в частности, процесс окислительного фосфорилирования на первом этапе работы ДЦ. Учитывая, что накопление витамина Е в печени кур сопровождалось увеличением концентрации витамина А, то стимуляция окисления 2-оксоглутарата может быть проявлением межвитаминных взаимодействий и связана как с действием витамина Е, так и влиянием витамина А на I комплекс дыхательной цепи.
Выводы:
1. Взаимодействия витаминов А и Е при одновременном длительном применении их в рационе кур проявляются в дозозависимом влиянии на энергетические процессы в митохондриях печени.
2. Совместное применение в рационе кур-несушек витамина Е, в количестве 100 мг/кг и витамина А — 10 тыс. МЕ/кг корма на протяжении 40 дней, оказывает стимулирующее влияние на реакции тканевого дыхания в печени кур и создаёт возможность обогащения яиц этими витаминами.
3. Витамины А и Е, накапливаясь в печени цыплят в результате потребления их курами-несушками в повышенных дозах (витамин А — 100 и 400 тыс. МЕ/кг, витамин Е — 200 мг/кг), оказывают негативное влияние на энергетические процессы у потомства.
Список литературы:
1.Витамины в питании животных / А.Р. Вальдман, П.Ф. Сурай, И.А. Ионов, Н.И. Сахацкий. Харьков: РИП Оригинал, 1993. — 423 с.
2.Донченко Г.В. Молекулярные механизмы влияния витамина Е на тканевое дыхание и энергетический обмен // Труды V Всесоюзного биохимического съезда. М., — 1985. — Том 3. — С. 48—49.
3.Ионов И.А. Обогащение яиц и мяса птицы витамином Е // Вісник аграрної науки. — 1997. — № 10. — С. 48—51.
4.Карабанов С.Е. Нормы кормления птицы различных видовых и половозрастных групп. [Электронный ресурс] — Режим доступа. — URL: http://poultry-new.narod.ru/DOC/Doc1.htm#1. (дата обращения: 29.01.2014).
5.Критерии и методы контроля метаболизма в организме животных и птиц: справочное пособие [Под. ред. Ионов И.А.]. Харьков: Ин-т животноводства НААН, 2011. — 377 с.
6.Лемешева М.М. Кормление сельскохозяйственной птицы. Сумы: Слобожанщина, 2003. — 152 с.
7.The cell death regulator GRIM-19 is an inhibitor of signal transducer and activator of transcription / J. Zhang, J. Yang, S.K. Roy [at all.] // Cell Biology. — 2003. — V. 100, — № 16. — P. 9342—9347.
8.Higdon J. Vitamin E [Electronic Resource] / J. Higdon, V.J. Drake, M.G. Traber // Linus Pauling Institute, Micronutrient Information Center, Oregon State University. — Copyright 2008. — Mode of access: [Электронный ресурс] — Режим доступа. — URL: http://lpi.oregonstate.edu/infocenter/vitamins/vitaminE/ Title from the screen (дата обращения: 23.01.2014).
9.Vitamin A [Electronic Resource] / J. Higdon, V.J. Drake, J. Mayer / [at all.] // Linus Pauling Institute, Oregon State University, Micronutrient Information Center. — Copyright 2000-2014. — Mode of access: [Электронный ресурс] — Режим доступа. — URL: http://lpi.oregonstate.edu/infocenter/vitamins/vitaminA/. – Title from the screen (дата обращения: 29.01.2014).
дипломов