
Статья опубликована в рамках: IV Международной научно-практической конференции «Научные достижения биологии, химии, физики» (Россия, г. Новосибирск, 01 февраля 2012 г.)
Наука: Биология
Секция: Ботаника
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов
АНАЛИЗ РАСПОЛОЖЕНИЯ ЦВЕТКОВ НА РАСТЕНИЯХ НА ОСНОВАНИИ ГОМОЛОГИЧНЫХ МОРФОЛОГИЧЕСКИХ БЛОКОВ
Харченко Виктория Евгеньевна
канд. биол. наук, доцент кафедры биологии растений, Луганский национальный аграрный университет, г. Луганск, Украина
Расположение цветков имеетважное адаптивное значение для Angiospermae, так как обуславливает эффективность воспроизведения растений [9]. На генеративных побегах растений может формироваться одиночный цветок или упорядоченное множество цветков — соцветие. В ходе длительной истории изучения расположения цветков на растениях, термин «соцветие», так и не получил чёткого определёния. Это связано с тем, что расположение цветков на растениях, может изменяться в ходе морфогенеза и под влиянием взаимодействия генотипа с условиями среды. Для анализа расположения цветков на растениях, сформировалось несколько разноплановых подходов, которые не всегда охватывали одинаковый круг вопросов. В частности, К. Линней (1751), анализировал расположение цветков, J. D. Hooker (1858), A. W.Eichler (1875),K. Goebel (1889)—цветков и цветоносных ветвей, W. Troll (1934) — цветоносных осей [5, 10, 14, 16, 27, 28]. Это привело к разночтениям в трактовке терминов и проблемам в классификации соцветий. На недоработки основополагающих вопросов по определению принципов классификации и разграничения соцветия и вегетативной сферы указывали в своих работах многие морфологи: A. W. Eichler (1875), C. Nägeli (1855), J. Velonovsky (1910), H. W. Rickett (1944), F. Schroeder (1987), H. Gaussen (1952), H.-J. Maresquelle (1964), Y. Sell (1964), Т. В. Кузнецова (1991)и др. [3, 13, 14, 19, 21, 23, 26, 29]. В частности H. W. Rickett, отмечал, что в терминах и понятиях морфологии соцветий существует беспорядок, который не может быть решён по принципу приоритета, однако в процессе обсуждения терминологии могут быть выявлены проблемы, которые ранее не были обозначены и нуждаются в исследовании [24].
При анализе расположения цветков у растений, по мнению J. Roeper (1826), G. Bentman, J. D. Hooker (1918),W. Troll (1964) и др. принципиальное значение имеет верхушечный цветок [12, 25, 27]. Согласно современным представлениям, к анализу расположения цветков на растениях, сформулированных в работах H. Gaussen,H.-J. Maresquelle, Y. Sell, Т. В. Кузнецовой, на побеге формируется флоральная единица, которая может быть представлена одиночным цветком или соцветием (простым, сложным, составным), в зависимости от взаимодействия генотипа с условиями среды [4, 15, 19]. Таким образом, одиночный цветок на верхушке побега может представлять собой и самостоятельную флоральную единицу и её часть.
Тело растения имеет модульную организацию, а его нарастание происходит в течение всей жизни, с чередующимися циклами роста и покоя [17]. Это приводит к образованию гомологичных морфологических блоков. При морфологическом описании растения, согласно работам С. В. Мейена (1978), начальной задачей является расчленение его на мероны (произвольно выделенные части растения — модули), а второй — реконструкция из полученных меронов исходного образа [7, 20]. Одну и ту же систему можно расчленить на элементы бесконечным количеством способов и сформулировать бесконечное количество алгоритмов сборки. Проблема состоит в том, чтобы расчленить тело растения на возможно более естественные части с точки зрения природы растительного организма и найти правила композиции, отражающие природные процессы формирования из этих элементов целостной системы [3]. Процесс выделения модулей в пределах одного организма можно расценивать как процедуру сериальной гомологизации [1].
Для компьютерного моделирования развития растений в работах A. Lindenmayer, P. Prusinkiewicz была использована теория фракталов Б. Мандельборта [6, 18, 22]. Однако структуры компьютерных моделей растения и реальных растений не всегда идентичны. Это связано, с тем, что морфологические блоки, выделенные на растении не всегда гомологичные, а реальное развитие растений может быть смоделировано только на основе гомологичных морфологических блоков (модуль) [7]. Морфологические блоки, выделенные на растении, естественны в том случае если, подобно фракталам, обладают свойствами самовоспроизводящихся структур, то есть в процессе развития растения воспроизводятся на нём с уменьшением масштабов. Применение к растениям системы модулей состоящими из гомологичных элементов позволяет избежать излишней детализации и произвольных трактовок составляющих элементов, которые затрудняют анализ расположения цветков на растениях. По мнению С. В. Мейена, если понимать под гомологизацией классификацию частей (компонентов, элементов, модулей, меронов), то анализ модулей в качестве конструктивных частей тела растений расширяет возможности сравнительной морфологии [5]. Сравнение модулей разного иерархического уровня позволяет выявить конструктивные особенности органов растений в филогенетически далеких таксонах, понять механизмы возникновения конвергенций и параллелизмов [8]. В связи с этим, на основании поэтапных сопоставления онтогенеза (морфогенеза) растений по филогенетической линии могут быть выявлены модусы трансформации морфологических блоков и составляющих их элементов
В основе морфологии растений и кибернетики находятся сходные принципы. В частности фундаментальным понятием кибернетики, W. R. Ashbу (1956) считал, понятие различия, то есть различимы два объекта по существу, либо один объект изменяется во времени [10]. Это так же справедливо и для морфологии растений. Во-первых, потому, что метамерия растений иерархична и представляет собой систему соподчиненных метамеров разного иерархического уровня [1]. Во-вторых, потому, что побеги растенийобладают свойствами самовоспроизводящихся структур, то есть со временем, в процессе развития растения, воспроизводятся на нём с уменьшением масштабов подобно фракталам.
Все элементы растения могут изменяться или редуцироваться согласовано и независимо от других [31]. Поэтому вариантов трансформации структуры побегов может быть бесконечное множество, следовательно, целесообразно установить не столько все возможные варианты расположения элементов побегов, сколько определить модусы их преобразования и возможные ретроспективы в пределах изучаемого таксона или территории.
Учитывая, что у растений наблюдается смена фаз развития их можно рассматривать как динамичную структуру, которая претерпевает изменения поэтапно — подобно фракталу. Системы воспроизведения модулей на растении подвержены влиянию факторов внутреннего и внешнего происхождения, следовательно, их состав и структура могут варьировать. Граница между разными морфологическими блоками может быть установлена на основании сопоставления растений на разных фазах морфогенеза (до и после перехода к цветению). Поэтому морфологический анализ расположения цветков на растении целесообразно проводить поэтапно.
В связи с тем, что в ряде случаев трактовка терминов для описания расположения цветков на растениях может быть спорной, для обозначения выделенных на растении морфологических блоков (модулей) целесообразно использовать минонимы, как это было предложено Т. В. Кузнецовой: флоральная единица — ФЕ [4].
При модульном анализе расположения цветков всё растение расценивается как общее множество — булеан, в составе которого имеются соподчинённые подмножества. Под влиянием факторов внутреннего и внешнего происхождения состав множеств и подмножеств может варьировать ([1; +∞)). Таким образом, побег растения можно представить в виде нескольких нечётких множеств (обобщённых в смысле Гогена) [30]. При переходе к цветению на побеге развивается флоральная единица которая может быть представлена одним цветком или множеством цветков — соцветием. Под влиянием факторов различного происхождения число цветков в составе флоральной единицы может варьировать. В контексте термина «соцветие» предполагает наличие нескольких цветков, но по ряду причин, они могут остаться недоразвитыми, а в случае если сформируется только один цветок, использование термина «соцветие» становится не вполне корректным. Под «флоральной единицей» равнозначно можно понимать: один цветок, простое, сложное или составное соцветие. Поэтому термин«флоральная единица» предпочтительней чем «соцветие», а при разграничении растения на модули он может расцениваться как элементарный модуль (Мodulus elementarius —ME). При поэтапном морфологическом анализе структура и состав морфологического блока другого уровня не имеет значения. Флоральная единица (ME) является соподчинённым ему элементом побега, и может быть охарактеризована как исходный модуль (Modulus initialis —MI). Соцветия состоят из метамеров, которые могут образовывать дублирующиеся блоки — модули метамеров (Modulus metamus —MM). Таким образом, побег (MI) можно рассматривать как исходное множество, в составе которого имеется подмножество — флоральная единица (ME), состоящая из подмножеств — метамеров соцветия (MM).
На начальном этапе морфологического анализа расположения цветков на растении представляется целесообразным определение структуры и состава побегов (MI) главного и боковых, а затем границы флоральной единицы (ME). В связи с тем, что при поэтапном морфологическом анализе структура и состав морфологического блока (модуля) другого уровня не имеет значения на этапе анализа состава и расположения побегов (MI) структура и состав флоральных единиц (ME) безразличны.
Побеги (MI) растения имеют разный уровень соподчинения, который обусловлен физиологически. Всё растение можно охарактеризовать как общий побег (communis —cMI), на котором имеется главный (primarialis —pMI) и боковые (lateralis —lMI), которые в зависимости от уровня ветвления могут быть первого порядка (1-lMI), второго (2-lMI) и т. д. (рис. 1).
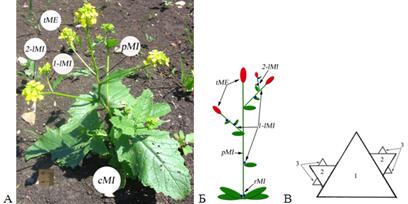
Рисунок 1. Модульный анализ расположения цветков на растении Sinapis arvensis
А – сMI - общий побег S. arvensis: pMI — главный побег, 1-lMI — боковые побеги первого порядка(1-lMI), 2-lMI — боковые побеги второго порядка, tME — соцветие на верхушке побега; Б — схема растения: pMI– главный побег, 1-lMI — боковые побеги первого порядка(1-lMI), 2-lMI — боковые побеги второго порядка, tME — соцветие на верхушке побега, 1-rMI — зачаток розеточного побега, В — схема фрактального треугольника построенного по принципу ветвления побега S. arvensis: 1 — главный побег (pMI), 2 — боковой побег (1-lMI), 3 — боковой побег второго порядка (2-lMI). Схематические обозначения:  — лист,
— лист,  — флоральная единица (ME),
— флоральная единица (ME),  — зачаток бокового побега (lMI),
— зачаток бокового побега (lMI),  — исходный модуль (MI);
— исходный модуль (MI);
В случае, когда часть листьев побега собрана в прикорневую розетку, боковые побеги, которые развиваются из их пазухи, могут быть охарактеризованы как розеточные (rosulata — rMI), так как это указывает на неравномерное развитие междоузлий, что является существенным признаком, при описании структуры побега.
В состав побега (MI) Sinapis arvensis входят: лист, междоузлие и флоральная единица (ME) (рис.1 В). В пазухах листьев находятся зачатки таких же морфологических блоков (lMI), которые в зависимости от внешних условий могут развиться в большей или меньшей степени. Структурная иерархия побега Sinapis arvensis (рис. 1 А) может быть проиллюстрирована при помощи фрактального треугольника (рис. 1 В).
Состав, расположение и модус воспроизведения модулей обусловлены внутренними факторами (генотипом), а уровень иерархии (ветвления) ограничен внешними факторами (условиями среды). Поэтому состав и структура метамеров побегов и соцветий могут варьировать. Пределы варьирования состава и структуры морфологических блоков могут быть установлены на основании сопоставления модификационной и мутационной изменчивости у филогенетически близких таксонов.
Так как клетки меристемы тотипатентны [11], а специализация побега обусловлена гормонально [2], флоральные единицы могут формироваться из меристемы разной локализации: апикальной (Adoxa moschatellina (Adoxaceae), Erophila verna(Brassicaceae), Tulipa quercetorum (Liliaceae), Delphinium puniceum (Ranunculaceae), Lythrum salicaria (Lythraceae), Utricularia vulgaris (Lentibulariaceae), Althaea officinalis (Althaea officinalis), Plantago major (Plantaginaceae) и др.), пазушной (Hippuris vulgaris (Hippuridaceae) и др.), вставочной (HippophaerhamnoidesL.(Elaeagnaceae)) ираневой (Córnus mas (Соrnасеае)) —каулифлоия.
Цветки, листья и прилистники могут иметь разный генезис, то есть они могу принадлежать верхушечной флоральной единице или боковому побегу.
Боковые побеги закладываются в пазухах листьев, поэтому их расположение соответствует филлотаксису растения (рис. 2).
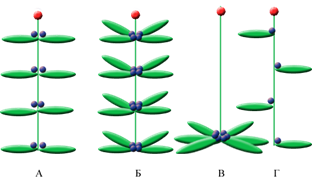
Рисунок 2. Схемы расположения на побегах зачатков флоральных единиц (ME) и боковых побегов, у растений с различным филотаксисом
А — супротивное листорасположение (Adoxa moschatellina (Adoxaceae)),
Б — мутовчатое листорасположение (Utricularia vulgaris (Lentibulariaceae)),
В — розеточное листорасположение (Erophila verna(Brassicaceae)),
Г — очередное листорасположение (Matthiola longipetala (Brassicaceae)); Схематические обозначения:  — лист;
— лист;  — зачаток бокового побега;
— зачаток бокового побега;  — зачаток флоральной единицы (ME).
— зачаток флоральной единицы (ME).
Листья дифференцируются из примордиев апикальной меристемы, поэтому пазушная меристема листьев — это остаток апикальной меристемы. Таким образом, участки пазушной меристемы закладываются по направлению сверху вниз и активизируются сходным образом (рис. 2). Флоральные единицы, чаще всего, занимают верхушечное положение на побегах. В их составе формируются прицветники в пазухах, которых образуются цветки или боковые оси соцветия, то есть боковые флоральные единицы. Изменение расположение прицветников в составе флоральной единицы, по сравнению с листорасположением побега, обусловленное переходом к цветению предполагает так же сходное изменение расположения цветков. В связи с тем, что элементы флоральной единицы (ME) формируются по направлению снизу вверх граница между флоральными единицами (ME) и боковыми побегами (lMI) может быть установлена на основании сопоставления морфологических блоков растения до и после перехода к цветению.
После установления на побеге границ флоральной единицы целесообразно проанализировать расположения цветков её составе. Для характеристики структуры соцветия существенное значение имеют: интенсивность ветвления его осей, соотношение длины междоузлий и цветоножек и расположение цветков на оси соцветия. В зависимости от интенсивности ветвления оси соцветия подразделяют на простые (simplex — sME) и сложные(compositus —cME) [14]. У сложных соцветий от главной оси (primaria — pME), отходят боковые оси (lateralis —lME), на которых формируются боковые соцветия первого порядка (1-lME), второго порядка (2-lME) и т. д. Ветвление соцветий может быть однородным (cME) (например кисть из кистей)или разнородным (например, кисть из корзинок), в таком случае соцветие характеризуют как составное (агрегатное)(aggregatae — aME) (рис. 3).

Рисунок 3. Иерархия осей соцветия
А — простое соцветие (sME) Sinapis arvensis (Brassicaceae), Б — сложное соцветие (cME) Aesculus hippocastanum (Hippocastanaceae): pME — соцветие на главной оси —тирс (кисть), 1-lME — боковые соцветия —кисти; В — составное соцветие (aME) Ambrosiatrifida(Asteraceae): pME — соцветие на главной оси —кисть, 1-lME — боковые соцветия 1-го порядка —корзинки.
В связи с тем, что сходные морфологические признаки могут иметь разное происхождение, для установления их генезиса, целесообразно проведение сопоставлений гомологичных морфологических блоков растений у филогенетически близких таксонов.
При определении состава и границы соцветия на побеге целесообразно учитывать ритмологические особенности формирования цветков на верхушке растения и на боковых побегах, а так же генезис составляющих его элементов. В отличие от побега (MI) в состав, которого могут входить: стебель, листья, прилистники и флоральная единица (ME), в пазухах листьев соцветия формируются не боковые побеги, а цветки или соцветия. Состав побега и флоральной единицы может варьировать. Цветок в пазухе листа может быть: составным элементом облиственного соцветия, боковым побегом (состав которого редуцирован до одного цветка), элементом боковой оси сложного соцветия (недоразвитым на данном этапе морфогенеза). Таким образом, один и тот же морфологический элемент может иметь разное происхождение. Генезис признака определяет ретроспективу его преобразований в ходе эволюции таксона, поэтому имеет принципиальное значение при морфологическом анализе растений.
Эволюционные преобразования (аббревиация, пролонгация, девиация и др.) могут быть направлены на преобразование модулей разного уровня иерархии растения: побега (MI), флоральной единицы (ME) или блоков метамеров флоральных единиц (MM). В частности, в ходе эволюции таксонов у одних растений может наблюдаться девиация расположения побегов (MI), положительная анаболия в составе флоральных единиц (ME) и отрицательная анаболия в составе метамеров соцветия (MM). Поэтому для установления эволюционных тенденций у таксонов целесообразно проводить филогенентические сопоставления поэтапно с учётом уровня иерархии морфологических блоков и стадий морфогенеза.
Список литературы:
1. Гатцук Л. Е. Растительный организм: опыт построения иерархической системы его структурно-биологических единиц // Современные подходы к описанию структуры растений. 2008. — С. 26 — 47.
2. Кине Ж.-М., Сакс Р., Бернье Ж. Физиология цветения Том 3. Развитие цветков. М.: 1991. — 445 с.
3. Корона В. В. Основы структурного анализа в морфологии растений. Свердловск: 1987. — 272 с.
4. Кузнецова Т. В. Морфология соцветий: современное состояние / Кузнецова Т. В. // ВИНИТИ. Итоги науки и техники. Ботаника. Т. 12 Современные методы систематики высших растений. Морфология и экология репродуктивных органов цветковых растений. М.: 1991. — С. 51—124.
5. Линней К. Философия ботаники. М.: 1989. — 456 с.
6. Мандельброт Б. Фрактальная геометрия природы М.: 2002. — 656 с.
7. Мейен С. В. Основные аспекты типологии организмов // Журнал общей биологии. 1978. Т. 39. № 4. — С. 495 — 508
8. Нотов А. А. Модульные организмы как объекты исследования в систематике и морфологии // Карл Линней в России. Философский век: альманах: СПб.: 2007. — С. 187 — 189.
9. Тахтаджян А. Л. Вопросы эволюционной морфологии растений/ Л., 1964. — 214 с.
10. Ashbу W. R. An Introduction to Cybernetics. London: 1956. — 432 p.
11. Barlow P. W. Meristems, metamers and modules and the development of shoot and root systems // Bot. J. Linn. Soc. 1989. № 100. — P. 255 — 287.
12. Bentman G., Hooker J. D. The flowering plants and ferns Londini: L. Reeve and Co., LTD. 1918. — 584 p.
13. Cerceau-Larrival M.-T. H., Gaussen M. H. Morphologie des cotyledons d'Ombelliferes II Comptes rendus hebdomadaires des seances de Tacademie des seances / 1957. T. 244. — № 5. — P. 650—660.
14. Eichler A. W. Bluetendiagramme 1. Leipzig: 1875. 345 p.
15. Gaussen H. L'evolution pseudocyclique // Année biologique1952. Ser.3. Т. 28. Fasc. 7 — 8. P. 207 — 225.
16. Goebel K. Blutenbildung und Sprossgestaltung (Anthokladien und Infloreszenzen) 2. Ergdnzungsband zur 'Organographie der Pflanzen'. Jena. 1931. — 270 p. Режим доступа: http://www.archive.org/stream
17. Jackson J. B. C., Coates A. G. Life cycles and evolution of clonal (modular) animals // Phil. Trans. R. Soc. London: 1986. — Ser. B.313. — P. 7 — 22.
18. Lindenmayer A., Prusinkiewicz P. Developmental models of multicellular organisms а computer graphics perspective // In C. Langton, editor, Artificial Life: Proceedings of an Interdisciplinary Workshop on the Synthesis and Simulation of Living Systems held September, in Los Alamos, New Mexico, 1987. — P. 221—249.
19. Maresquelle H.-J., Sell Y. Les problemes physiologiques de la floraison descendante // Bull, de la Societe franais de la physiologie vegetale. 1965. № 11(1). — P. 94 — 98.
20. Meyen S. V. Plant morphology in its nomothetical aspects // Bot. Rev. 1973. — Vol. 39. № 3. — P. 205 — 260.
21. Nägeli C., Cramer C. Pflanzenphysiologische Untersuchungen Heft. Zürich. 1855. V1. —120 p.
22. Prusinkiewicz P., Erasmus Y., Lane B., Hareder L. D., Coen E. Evolution and development of inflorescence architectures // Science. 2007. V. 316. no.5830 — P. 1452—1456.
23. Rickett H. W. Inflorescence of Philadelphus // Amer. Midl. Nat. 1944. № 31. —P. 759—762.
24. Rickett H. W. Materials for dictionary of botanical terms — III Inflorescence // Bulletin of the Torrey Botanical Club., 1955. — V.82(6). — P. 419— 445.
25. Roeper J. Observationes aliquot in florum inflorescentiarum que naturam // Linnaea 1. 1826. — P. 433 — 466.
26. Schroeder F. G. Infloreszenzen, Synfloreszenzen und Moduln. Eine terminologische Beitrag zur Infloreszenzmorphologie // Ibid. 1987. — Bd. 108. — Hf. 2/3. — S. 449—471.
27. Troll W. Die Infloreszenzen Bd. 1. Jena, 1964. — 615 s.
28. Troll W. Die Infloreszenzen Bd. 2 Veb Gustav Fischer Verlag. Jena, 1969. —630 s.
29. Velonovsky J. Vergleichende Morphologie der Pflanzen // III Teil. Prag; Rivnac. 1910. — S. 733— 1211
30. Zadeh L. Fuzzy sets // Information and Control. 1965. v.13 (8). — P. 338—353.
31. Zimmermann W. Strenge Objekt Subjekt—Scheidung als Voraussetzung wissenschaftlicher Biologie// Annalen der Philosophie. 1937. № 15. — P. 1—44.
дипломов