Статья опубликована в рамках: III Международной научно-практической конференции «Научные достижения биологии, химии, физики» (Россия, г. Новосибирск, 27 декабря 2011 г.)
Наука: Биология
Секция: Физиология
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов
ХАРАКТЕР ВЛИЯНИЯ СЕЛЕКТИВНОГО СТИМУЛИРОВАНИЯ АКТИВНОСТИ МОНОАМИНЕРГИЧЕСКИХ НЕЙРОМЕДИАТОРНЫХ СИСТЕМ МОЗГА НА ПСИХОДИНАМИЧЕСКИЕ ПОКАЗАТЕЛИ ПОВЕДЕНИЯ САМЦОВ БЕЛЫХ КРЫС
Фролова Галина Александровна
канд. биол. наук, ст. преподаватель ДонНУ, г. Донецк
E-mail: gljukkk@ukr.net
Необходимостьисследования механизмов возникновения психической или поведенческой депрессии связано со значительным увеличением количества людей, страдающих депрессивные расстройства. Высокая частота возникновения пограничных нервно-психических расстройств обусловлена ростом эмоционального и информационной нагрузки в условиях ускоренного научно-технического прогресса и темпа жизни в целом. Об актуальности проблемы депрессий свидетельствуют эпидемиологические данные: распространение расстройств депрессивного характера у населения стран Европы и США составляет сегодня не менее 5‑10%. Актуальность этой проблемы в общей медицине, где частота возникновения депрессий достигает 22‑33% и превышает такое распространенное заболевание, как артериальная гипертензия [13], существенно возрастает.
Насегодня наиболее разработанной является моноаминергических теория патогенеза депрессий, причем большинство исследователей считают, что при различных депрессивных состояниях могут нарушаться преимущественно или адренергические [8, 12], или серотонинергические механизмы [1, 5, 10], или оба вместе [2, 3, 6].
В последнее время появляются работы, результаты которых свидетельствуют о том, что в основе развития тех или иных депрессивных и тревожных расстройств может лежать не только угнетение активности вышеуказанных нейромедиаторных систем, но и их активация. В связи с этим, целью представленного фрагмента комплексной работы является анализ влияния селективной стимуляции активности моноаминергических нейромедиаторных систем на психодинамические показатели самцов белых крыс с исходно разным уровнем поведенческой активности.
Материалы и методы. Эксперимент был выполнен на 120 половозрелых беспородных крысах-самцах массой 180‑220 г, содержавшихся в стандартных условиях вивария. Эксперименты осуществлялись в светлое время суток. Для определения психодинамических характеристик у исследуемых животных использовали батарею следующих тестов: продырявленное поле, приподнятый крестообразный лабиринт, тест Порсолта.
На начальном этапе исследования исходная группа крыс по результатам тестирования в продырявленном поле была разделена на 3 подгруппы, согласно выраженности поведенческой активности. В последующем, прослеживали, каким образов фармакологические агенты, избирательно стимулирующие активность моноаминергических нейромедиаторных систем мозга влияют на проявления психодинамических характеристик подгрупп крыс (активности, тревожности, депрессивности), сформированных по результатам тестирования в продырявленном поле.
Уровень поведенческой активности устанавливали в продырявленного поле (ПП) согласно выражением исследовательской активности (ИА) [4]. Продырявленное поле представляет собой открытый пластиковый ящик 60×60×40 см., пол которого окрашена в зелено-голубой цвет и разделена линиями на 9 квадратов 20×20 см. По периметру квадратов в полу просверлены 12 отверстий диаметром 3 см. Продолжительность теста составляла 5 мин. Регистрировались такие поведенческие показатели: количество пересеченных квадратов (двигательная активность — ДА), стойки и количество обследованных отверстий (в сумме — исследовательская активность). Отдельно фиксировали количество фекальных болюсов, частоту уринации и количество актов груминга.
Уровень тревожности (УТ) определяли с помощью теста «приподнятый крестообразный лабиринт» (ПКЛ), регистрируя время пребывания животных на открытом пространстве лабиринта [7]. Лабиринт состоит из двух закрытых (огражденных с двух сторон) и двух открытых рукавов (10×50 см.), расположенных крестообразно на высоте 80 см. над уровнем пола. Тестирование начиналось с размещения экспериментального животного в центр лабиринта в положении носом в открытый рукав. В течение 5 минут проводилось наблюдение за поведением крысы. Регистрировались такие поведенческие акты: количество повторных выходов на открытую часть ПКЛ (открытые рукава + центральная платформа) и время пребывания в ней, количество визирань из закрытых рукавов, количество стоек на открытом пространстве и частота дефекаций.
В тесте Порсолта («вынужденного плавания», ВП) определяли уровень депрессивности животных [11]. При тестировании крыс на 6 мин. погружали в прозрачный пластиковый цилиндр высотой 60 см. и диаметром 50 см., наполненный водой (температура 27‑28°С), таким образом, чтобы животное не имело возможности опереться задними конечностями или хвостом о дно цилиндра. Поведенческими показателями служили: количество и время периодов полной недвижимости — индекс депрессивности (ИД). Для характеристики временной структуры вынужденного плавания подсчитывали количество и продолжительность периодов недвижимости, группируя их по четырем основным диапазонам: менее 6 секунд, от 6 до 18, от 18 до 36 и более 36 секунд. Учитывалась также количество фекальных болюсов.
По результатам тестирования был составлен поведенческий профиль популяции в исходных (контрольных) условиях. С целью нивелирования сезонных изменений в поведении исходная группа крыс (n=120) была разделена случайным образом на 3 подгруппы по 40 животных в каждой так, чтобы профиль каждой подгруппы отвечал исходном общем профиля. Таким образом, были образованы 3 однородных подгруппы, дальнейшая работа с которыми проводилась параллельно.
Выбор фармакологических препаратов осуществлялся согласно поставленными задачами с учетом точек применения их действия так, чтобы ожидаемый эффект был получен путем воздействия или на ферментную систему, участвует в процессе биологического синтеза нейромедиаторов, или на систему МАО, обеспечивающие разрушение моноаминов в синаптической щели.
Были использованы внутрибрюшные инъекции таких фармакологических препаратов: предшественника серотонина — L‑триптофана («Sigma», США) в дозе 100 мг/кг в течение 14 дней (Neume A. at al., 1999), селективного ингибитора обратного захвата норадреналина мапротилина («Sigma», USA) в дозе 20 мг/кг, в течение 14 дней (Guillem K. at al., 2006); специфического ингибитора МАО-Б селегилина (депренил, «Sigma», США) в дозе 3 мг/кг в течение 14 дней [3].
Первичныеэкспериментальные данные обрабатывались с помощью общепринятых методов математической статистики. Разделение исследуемой популяции животных на группы с разным уровнем поведенческой активности осуществлялось согласно правилу ±0,67δ. Для оценки достоверности различий между результатами контрольных исследований и для оценки достоверности различий между опытными и контрольными данными использовался U‑критерий Манна-Уитни. С помощью корреляционного анализа устанавливали наличие связей между определяемыми психодинамическими показателями животных. Математическая обработка материала осуществлялась с помощью пакета программ STATISTIСA 6.0 и Microcoft Excel.
Результаты и их обсуждение.Характер распределения исходной группы крыс на подгруппы по результатам контрольного тестирования в условиях продырявленного поля представлен на рисунке 1.
Психодинамический профиль исходной группы, составленный на основании контрольного тестирования, в условиях используемой батареи тестов отражен в таблице 1.
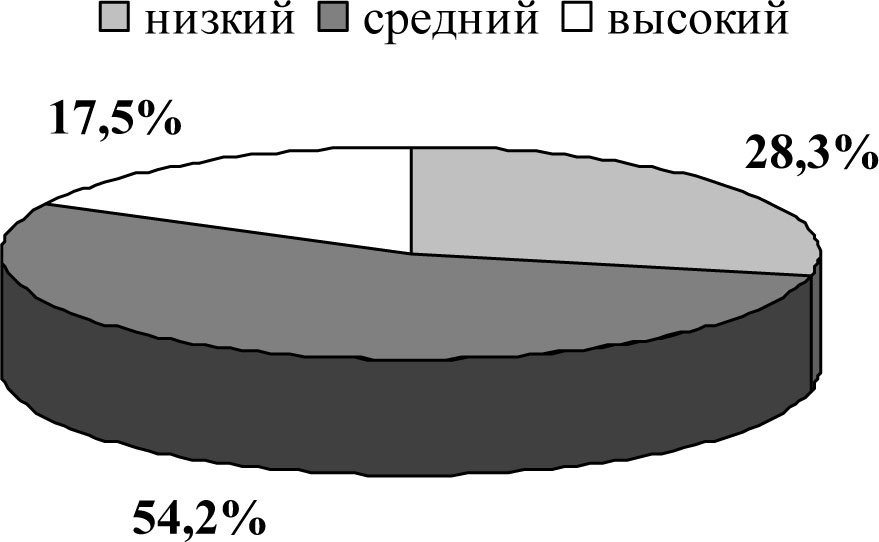
Рисунок 1. Распределение исходной группы животных (n=120) на подгруппы с разным уровнем поведенческой активности в условиях продырявленного поля.
Из таблицы следует, что животные с исходно средним и высоким уровнями выраженности исследовательской активности достоверно не отличались по уровню двигательной активности. Кроме того, исходно низкоактивные животные обладают максимальным уровнем тревожности, а исходно высокоактивные в продырявленном поле —минимальным показателем депрессивности, среднеактивные —минимальной суммой замираний в тесте Порсолта.
Анализ общегрупповых корреляционных связей показал, что исследовательская и двигательная активности связаны между собой корреляционной связью с коэффициентов rоб/кон=0,822 (p<0,01), кроме того, установлено, что показатель эмоциональности (количество фекальных болюсов), связан положительно с двигательной активностью (rоб/кон=0,245, p<0,01), временем пребывания на открытых рукавах ПЛК (rоб/кон=0,302, p<0,01), количеством замираний длительностью менее 6 секунд в тесте Порсолта (rоб/кон=0,217, p<0,05) и отрицательно с общим количеством замираний в ВП (rоб/кон=-0,446, p<0,05). Показатель депрессивности отрицательно связан с двигательной (rоб/кон=-0,321, p<0,01) и исследовательской (rоб/кон=-0,351, p<0,01) активностями и положительно с общим количеством замираний в тесте Порсолта (rоб/кон=0,219, p<0,05). Двигательная активность коррелирует с временем пребывания животных на открытом пространстве ПКЛ (rоб/кон=0,187, p<0,05). Отсутствие связей между некоторыми показателями тех или иных психодинамических характеристик указывает на участие в их контроле различных нейроанатомических образований и нейрохимических систем мозга.
Таблица 1.
Психодинамический профиль исследуемой популяции (n=120)
в контрольных условиях, ()
|
Показатели поведения |
Уровни поведенческой активности |
||
|
низкий (n=34) |
средний(n=65) |
высокий(n=21) |
|
|
Результаты тестирования в продырявленном поле |
|||
|
Исследовательская активность |
4,7±0,52●● |
17,0±0,45 |
29,4±1,04●●## |
|
Двигательная активность |
7,0±0,89●● |
25,9±1,10 |
37,1±1,21# |
|
Результаты тестирования в приподнятом крестообразном лабиринте |
|||
|
Время пребывания на открытом пространстве лабиринта, с. |
103,9±7,77● |
150,1±8,09 |
121,6±11,92 |
|
Результаты тестирования в тесте Порсолта |
|||
|
Суммарное время неподвижности, с. |
161,2±13,93 |
130,2±0,81 |
97,1±14,75# |
|
Общее количество периодов замираний |
13,9±0,74●● |
7,2±0,60 |
11,5±1,02● |
|
Количество замираний длительностью менее 6 секунд |
8,7±0,65 |
110,1±0,43 |
8,7±0,72 |
|
Количество фекальных болюсов |
4,0±0,33● |
5,4±0,24 |
4,4±0,45 |
Примечание: #, ## — различия статистически значимы (рu<0,05) и (рu<0,01) соответственно при сравнении показателей группы с крайними типами выраженности показателей активности.
●, ●● — различия статистически значимы (рu<0,05) и (рu<0,01) соответственно в сравнении показателей условного контроля (средний уровень активности) с группами высокого и низкого уровня показателей исследовательской активности.
Обращает на себя внимание, что при рассмотрении корреляционных связей показателей внутри каждой из выделенных подгрупп (т. е. с низкой, высокой и средней активностью), направленность некоторых зависимостей отличалась от общегрупповой (табл. 2). Прежде всего, обращает на себя внимание отсутствие корреляционных зависимостей у животных с исходно высоким уровнем активности в тех случаях, когда эти связи присутствовали при проведении анализа у всех животных без разделения на подгруппы. Так, в данной подгруппе крыс отсутствует связь между двигательной активностью такими показателями как: исследовательская активность, эмоциональность и суммарное количество замираний (см. табл. 2). Однако, обратная картина (присутствие зависимости у высокоактивных крыс на фоне ее отсутствия в общей группе животных) наблюдается между показателями коротких замираний в тесте Порсолта и общего их количества.
Селективное стимулирование активности нейромедиаторных систем по-разному повлияло на исследуемые психодинамические показатели белых крыс в пределах выделенных подгрупп.
Во-первых, установлено, что исследовательская и двигательная активность под воздействие выбранных фармакологических агентов изменяются однонаправлено: в случае стимулирования серотонинергической и норадренергической систем L-триптофаном и мапротилином, соответственно, наблюдалось угнетение выраженности данных показателей (рис. 2), а в случае стимуляции дофаминергической системы депренилом у исходно низко- и среднеактивных животных наблюдается рост как ДА (на 111,0±2,25 и 86,6±3,67% соответственно), так и ИА (на 79,1±2,45 и 52,4±3,97% соответственно), а у исходно высокоактивных — угнетение выраженности этих показателей.
Относительно изменений тревожности, то следует отметить анксиогенное влияния стимулирования норадренергической системы на крыс с исходно средним уровнем активности (рис. 3, А), поскольку суммарное время пребывания на открытом пространстве у этой подгруппы животных сократилось на 35,8±2,13% (pu<0,05). Депренил и L-триптофан, в основном, оказали анксиолитическое влияние на исследуемых животных (т. е. уровень тревожности снизился).
Таблица 2.
Корреляционные зависимости основных показателей у животных с разным уровнем поведенческой активности
|
Показатели |
Уровень активности |
Продырявленное поле |
Приподнятый крестообразный лабиринт |
Тест Порсолта |
||||
|
Активность |
Показатель депрессивности |
Количество замираний |
||||||
|
исследова- тельская |
двига- тельная |
Время пребывания на открытом пространстве |
коротких |
общее |
||||
|
Двигательная активность |
низкий |
0,687** |
|
|
|
|
|
|
|
средний |
0,501** |
|
|
|
|
|
||
|
высокий |
0,355 |
|
|
|
|
|
||
|
Время пребывания на открытом пространстве |
низкий |
0,397** |
0,310* |
|
|
|
|
|
|
средний |
0,054 |
0,011 |
|
|
|
|
||
|
высокий |
–0,235 |
–0,069 |
|
|
|
|
||
|
Показатель депрессивности |
низкий |
–0,172 |
–0,100 |
0,189 |
|
|
|
|
|
средний |
–0,190 |
–0,048 |
0,211* |
|
|
|
||
|
высокий |
–0,058 |
–0,309 |
0,020 |
|
|
|
||
|
Количество замираний |
коротких |
низкий |
0,216 |
0,383** |
–0,099 |
–0,304* |
|
|
|
средний |
–0,038 |
0,032 |
0,146 |
0,163 |
|
|
||
|
высокий |
–0,123 |
–0,251 |
0,158 |
0,190 |
|
|
||
|
общее |
низкий |
–0,511** |
–0,499** |
–0,255 |
0,385** |
0,014 |
|
|
|
средний |
0,010 |
–0,434** |
–0,161 |
–0,214* |
0,196 |
|
||
|
высокий |
–0,053 |
–0,214 |
–0,063 |
0,638** |
0,439* |
|
||
|
Количество дефекаций |
низкий |
0,405** |
0,535** |
0,223 |
0,089 |
0,534** |
0,003 |
|
|
средний |
–0,109 |
0,273* |
0,219* |
0,201 |
0,032 |
–0,544** |
||
|
высокий |
–0,124 |
–0,073 |
0,252 |
–0,085 |
0,026 |
–0,365* |
||
Примечание: *, ** — корреляционная связь статистически значима при p<0,05 и, p<0,01 соответственно.

Рисунок 2. Характер изменения исследовательской (А) и двигательной (Б) активностей в тесте продырявленное поле относительно исходных значений (приняты за 0) в подгруппах животных с исходно низким, средним и высоким уровнями активности.
*, ** —различия статистически значимы при рu<0,05 и рu<0,01, соответственно.
Следует отметить, что избирательная стимуляция моноаминергических медиаторных систем мозга снизила индекс депрессивности во всех подгруппах крыс. При чем, в случае изменения активности серотонинергической и дофаминергической систем степень снижения ИД зависела от исходного уровня активности животных (рис. 3, Б): чем выше в исходных условиях был ИА, тем в большей степени снизилась депрессивность. При этом, мапротилин не повлиял на количество коротких периодов неподвижности в тесте Порсолта сократив, при этом, суммарное количество замираний в среднем на 40‑25% (pu<0,01), а стимуляция активности остальные нейромедиаторных систем повлияла на количество замираний длительностью менее 6 секунд, но не затронула общее число замираний. Так, стимуляция дофаминергической системы депренилом привела к увеличению количества коротких замираний в среднем на 30‑50% (pu<0,01), а введение L-триптофана —к сокращению данного показателя поведения в тесте Порсолта на 70‑60% (pu<0,01) у исходно средне- и высокоактивных крыс и увеличению на 42,9±3,12% (pu<0,05) у низкоактивных в контроле крыс.
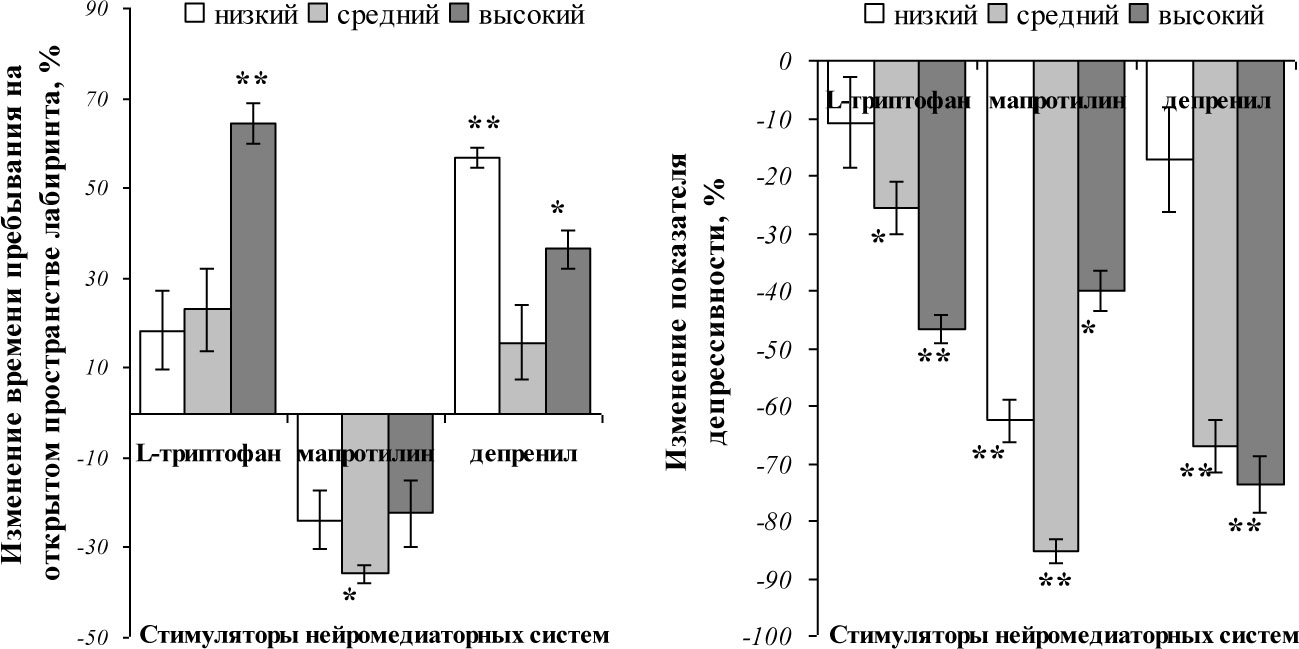
Рисунок 3. Характер изменения суммарного времени пребывания на открытом пространстве лабиринта (А) и индекса депрессивности (Б) в тесте Порсолта относительно исходных значений (приняты за 0) в подгруппах животных с исходно низким, средним и высоким уровнями активности.
*, ** —различия статистически значимы при рu<0,05 и рu<0,01, соответственно.
Обращает на себя внимание тот факт, что эмоциональность животных, оцениваемая по количеству фекальных болюсов, возросла у высокоактивных в контроле крыс при стимуляции серотонинергической (63,9±4,65, pu<0,01) и дофаминергической (48,5±3,66, pu<0,05) систем, что совпадает с характером изменения тревожности (ее снижения) у данных подгрупп крыс. У остальных самцов достоверных влияний на эмоциональность не выявлено.
При рассмотрении модификаций корреляционных связей на общегрупповом уровне после избирательного стимулирования активности нейромедиаторных систем мозга, обращает на себя внимание неизменность связи (p<0,01) между двигательной и исследовательской активностями (rоб/кон=0,822, rоб/L-трип=0,713, rоб/мапр=0,755, rоб/депр=0,512), в то время как по ряду других показателей корреляционные связи были утеряны. Интересным оказался тот факт, что после введение L-триптофана, мапротилина и депренила установились положительные корреляционные зависимости между количеством общих и коротких периодов замираний, в то время как в контроле эти показатели не были связаны между собой (rоб/кон=0,097, rоб/L-трип=0,940, rоб/мапр=0,722, rоб/депр=0,541).
Относительно корреляционных связей у животных с исходно разными уровнями исследовательской активности, то характер изменения таковых был настолько разнообразен и сложен, что их анализ выходит за пределы представленного фрагмента работы.
Таким образом, можно сделать выводы о том, что, характер влияния избирательного стимулирования активности серотонинергической и дофаминергической систем на психодинамические показатели самцов лабораторных животных находится в зависимости от исходного уровня поведенческой активности крыс. Эффекты стимуляции норадренергической системы мапротилином оказались более разнонаправленными.
Список литературы:
1.Августинович Д. Ф., Липина Т. В., Кудрявцева Н. Н. Особенности реакции серотонергической системы мозга на социальный стресс разной длительности у самцов мышей линий C57BL/6J и CBA/Lac // Рос. физиол. журн. им. И. В. Сеченова. 2001. Т. 87, № 4. С. 532‑542.
2.Андрух П. Г. Этиология, нейрохимия и клиника современных форм тревожно-депрессивных расстройств // Медицинские исследования. 2001. Т. 1, вып. 1. С. 109-110.
3.Исмайлова Х. Ю., Агаев Т. М., Семенова Т. П .Индивидуальные особенности поведения (моноаминергические механизмы) Баку: Нурлан, 2007. —229 с.
4.Калуев А. В. Стресс, тревожность и поведение: актуальные проблемы моделирования тревожного поведения у животных.Киев: CSF, 1998. —98 с.
5.Левада О. А. Нейробиология депрессии: серотониновая система мозга// Нейроnews. 2008. № 1. С. 25‑29.
6.Узбеков М. Г. Нейрохимимческие аспекты взаимодействия моноаминергических и гормональной систем в патогенезе тревожной депрессии // Социал. и клинич. психиатр. 2005. Т. 15, вып. 2. С. 108‑110.
7.Dawson G. R., Tricklebank M. D. Use of the elevated plus maze in the search for novel anxiolytic agents // Tr.in Pharmacolog.Sc. 1995. № 16. Р. 33‑36.
8.Fibiger H. C. Neurobiology of depression: focus on dopamine // Depression and Mania: From Neurobiology to Treatment. N. Y.: Raven Press, 1995.—216 р.
9.Kapur S., Mann J. J. Role of the dopaminergic system in depression // Biol. Psychiatry. 1992.Vol. 32,№ 1.P. 1‑17.
10.Mann, J. J., Role of the serotonergic system in the pathogenesis of major depression and suicidal behavior // Neuropsychopharmacology. 1999.№ 21.P. 99‑105.
11.Porsolt R. D., Pinchon M. L. Depression: a new animal model sensitive to antidepressant treatments // Nature. 1977. № 266. Р. 730‑732.
12.Ressier K. J., Nemeroff C. B. Role of serotoninergic noradrenergic system in the pathophysiology of depression and anxiety disorders // Depression and Anxiety. 2000. Vol. 12. Supl.1. P. 2‑19.
13Slade T, Andrews G. Generalized Anxiety Disorder and Major Depressive Disorder comorbidity in the National Survey of Mental Health and Well-Being // Depression and Anxiety. 2004.Vol. 20, № 1. P. 23‑31.
дипломов