
Статья опубликована в рамках: XXXI Международной научно-практической конференции «Инновации в науке» (Россия, г. Новосибирск, 31 марта 2014 г.)
Наука: Биология
Скачать книгу(-и): Сборник статей конференции, Сборник статей конференции часть II
- Условия публикаций
- Все статьи конференции
дипломов

Статья опубликована в рамках:
Выходные данные сборника:
АНАТОМИЧЕСКОЕ СТРОЕНИЕ «ВАЛЬКОВАТОГО ЛИСТА» CALLIGONUM APHYLLYM GÜRKE
Очиров Сергей Батнасанович
учитель химии и биологии МБОУ «Ики-Бурульская СОШ им. А. Пюрбеева», РФ, Республика Калмыкия, пос. Ики-Бурул.
E- mail: Ochirov_sergey@list.tu
Дорджиева Виктория Ильинична
канд. биол. наук, преподаватель кафедры ботаники и зоологии, доцент Калмыцкого Государственного Университета, РФ, Республика Калмыкия, г. Элиста
E- mail: Ochirov_sergey@list.tu
Манжиев Эренцен Юрьевич
студент 5 курса кафедры ботаники и зоологии Калмыцкого Государственного Университета, РФ, Республика Калмыкия, г. Элиста
E-mail :
ANATOMICAL STRUCTURE "TERETE LEAVES" CALLIGONUM APHYLLYM GÜRKE
Ochirov Sergey Batnasanovich
chemistry and biology teacher of MBOU “Iki-Burul school them. A. Purbeeva”, Russia Republic of Kalmykia Iki-Burul
Dordzhieva Victoria Ilinichna
cand. Biol. Sciences, lecturer in botany and zoology, associate Professor of Kalmyk State University, Russia Republic of Kalmykia Elista
Mangiev Erentsen Yur’evich
5th year student of the Department of botany and zoology of the Kalmyk State University, Russia Republic of Kalmykia Elista
АННОТАЦИЯ
Изучено морфолого-анатомическое строение «вальковатого листа» и побегов Calligonum aphyllym Gürke. Использован метод микроскопии и описание анатомических рисунков. Определены морфометрические показатели различных клеток листа.
ABSTRACT
The morphological and anatomical structure "terete sheet" and shoots Calligonum aphyllym Gürke were studied. Microscopy technique and description of anatomical drawings were used. Morphometric indices of various cells of the leaf were defined.
Ключевые слова : морфология; анатомия; эпидерма; вальковатый лист; побег.
Keywords: morphology; anatomy; epidermis; terete leaves; shoots.
Род Джузгун — Calligonum, из семейства Polygonaceae (Гречишные), представлен кустарниками или полукустарниками высотой около 30 см. Ксерофиты, адаптированные к условиям пустынь или полупустынь. Корни уходят в глубину до 30 метров, доходят до грунтовых вод; к тому же пески задерживают воду, то позволяет растению извлекать влагу уже на глубине 1,5 м. Виды джузгуна безлистны, а их короткие ассимиляционные побеги летом сбрасываются: веткопад в засуху [7]. Виды джузгуна активно используются как закрепители песков, разработаны способы создания долговременных плантации на песках [4—6]. В литературе имеются сведения по эволюции и систематике джузгуна [8, 10], рассматривается зависимость от условий среды [9]. Обращают внимание на формирование генеративной сферы. Отмечается высокая питательность, зеленые сочные веточки хорошо поедаются животными, особенно в жаркие дни [1, 2]. Сведения по анатомии вегетативных органов джузгуна отрывочны. В одном из учебных пособий приводится рисунок ветки и часть поперечного разреза через нее [3]. На рисунке показан лишь небольшой участок периферической части стебля с хлоренхимой и склеренхимными участками под эпидермой. Там же отмечено, «редукция листьев характерна для многих кустарников Средиземноморья, пустынь С. Азии и других мест с жарким и сухим климатом: джузгун, саксаул, испанский дрок, эфедра и многих других. У этих растений стебли принимают на себя функцию фотосинтеза, а листья или недоразвиваются, или опадают рано весной. В стеблях под эпидермой находится хорошо развитая палисадная ткань».
Материал и методика
Наиболее молодые гибкие фотосинтезирующие побеги Calligonum aphyllym Gürke (джузгун безлистный) были собраны на опытных участках по закреплению персков Черноземельского района РК в мае 2013 г. Анатомические срезы сделаны по общепринятой методике, рисунки — с микроскопа «Биолам» с использованием РА-4.
Результаты и обсуждения
Наиболее молодые верхушечные и пазушные «междоузлия» последнего порядка ветвления (заканчивающие рост побегов последнего года развития) по своей морфологии и анатомии больше напоминают вальковатые по форме, но удлиненные листья, чем завершающий безлистный побег междоузлие как принято считать. Завершающие побег «междоузлия» (верхушечные и пазушные) зеленые по окраске, гибкие, длиной, в условиях пустынь нашей республики, доходят до 20—27 мм, а в диаметре до 1,5 мм. Их верхушка тупозамкнута, а не завершена верхушечной точкой роста или почкой. И по анатомической структуре названные «междоузлия» побега представляют собой листья с проводящими пучками в центральной водозапасающей ткани (Рис. 1). В средней части конечных междоузлий молодых побегов, в водоносной паренхиме хорошо заметны три каллатеральных проводящих пучка. В остальных нижерасположенных междоузлиях молодого побега представлена сердцевина, вокруг которой в один круг расположены проводящие пучки, т. е. во всех остальных междоузлиях центральный цилиндр представлен типичной для представителей класса двудольных эустелой.
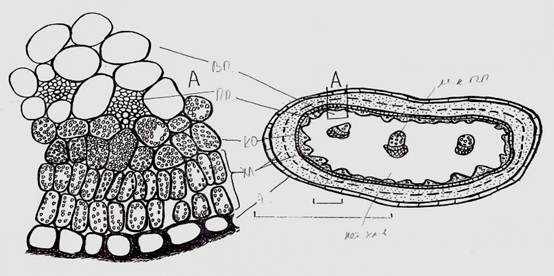
Рисунок 1. Схема и анатомическое строение (А) вальковатого листа C. aphyllym э. — эпидерма, хл. — хлоренхима, к.о. — кранц-обкладка, п.п. — проводящие пучки, в.п. — водозапасающая ткань, м.к.п.п. — медианный коллатеральный проводящий пучок
Отсутствие склеренхимных участков в хлоренхиме верхушечных и пазушных «междоузлий», и наличие таковых в хлоренхиме нижерасположенных междоузлий, является существенным структурным различием. Хорошо развитую хлоренхиму под эпидермой безлистных побегов отмечают у многих кустарников пустынь Средней Азии — джузгуна, саксаула, эфедры [7]. Однако, хлоренхима безлистного побега джузгуна составлена двумя слоями палисадных клеток и кранц-обкладкой из одного слоя почти квадратных по форме клеток. К кранц-обкладке конечных завершающих «междоузлий» по всей окружности подходят мелкие, почти чисто флоэмные, проводящие пучки. В некоторых из них заметны отдельные кольчатые сосуды, с очень узким диаметром, разбросанные без особого порядка (Рис. 1, А). Следует отметить, что число хлоропластов увеличивается от периферии к центру «междоузлия». В слое палисадных клеток прилегающих к эпидерме мало хлоропластов, во втором — много, а клетки кранц-обкладки буквально забиты хлоропластами. В пограничном прилегающем к кранц-обкладке клетках, хлоропласты встречаются только в верхней части «междоузлия», а уже ниже среднего уровня в этом пограничном слое остаются отдельные хлоропласты. Слой клеток кранц-обкладки остается забитым хлоропластами по всей длине «междоузлия» и намечает границу между хлоренхимой и центральной водозапасающей тканями. Отдельные хлоропласты можно обнаружить в периферийных клетках последней. В них же чаще всего накапливаются кристаллы солей (Рис. 2).
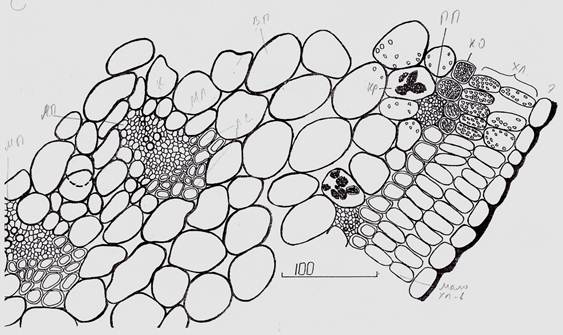
Рисунок 2. Анатомическое строение листа с медианным (м.п.) и латеральным (л.п.) проводящими пучками: кр.-кристаллы в клетках водозапасающей ткани, л.с. — лубяная склеренхима, м.л. — мягкий луб, к. — ксилема
В рассматриваемых, завершающих побег «междоузлиях» почти отсутствует механическая ткань. Исключение, три центральных поводящих пучка со стороны флоэмы несут слабо выраженную лубяную склеренхиму (Рис. 1, схема, Рис. 2). Стенки клеток лубяной склеренхимы настолько слабо выражены, что отличаются от окружающих клеток водозапасающей паренхимы только более мелкими размерами. В медианном проводящем пучке со стороны ксилемы представлена древесная склеренхима, которая не выражена в латеральных пучках. Ниже по «междоузлию» лубяная склеренхима трех центральных проводящих пучков уже четко представлена: типичная пяти-шестиугольная форма клеток с очень толстой оболочкой (Рис. 3).
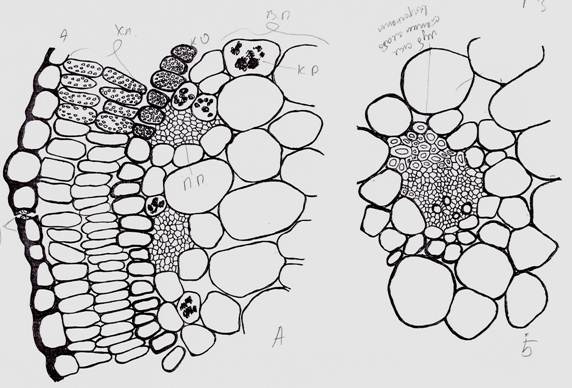
Рисунок 3. Анатомическое строение периферического участка вальковатого листа (А) и одного из латеральных проводящих пучков (Б) у. — устьица, п.п. — флоэмные проводящие пучки. Остальные обозначения как на рис. 1,2.
На эпидерме рассматриваемого конечного «междоузлия» хорошо представлены аномоцитные устьица, окруженные 4 (реже 3) околоустьичными клетками. Устьица на эпидерме ориентированы без особого порядка (Рис. 4). Число устьиц: 125 на 1 мм2.
Параметры отдельных тканей вальковатого листа внесены в табл. 1. Выше нами было отмечено возрастание числа хлоропластов от субэпидермального слоя палисадных клеток хлоренхимы к клеткам кранц-обкладки. Из табл. 1 видно, что клетки кранц-обкладки, буквально забитые хлоропластами, уступают палисадным клеткам по длине, по ширине же наоборот вытягиваются процентов на 30. Периферические проводящие пучки по своим параметрам в 3—4 раза меньше трех центральных. Высота эпидермальных клеток колеблется от 14 до 20 мкм, толщина кутикулы достигает до 30—40 % высоты эпидермы.
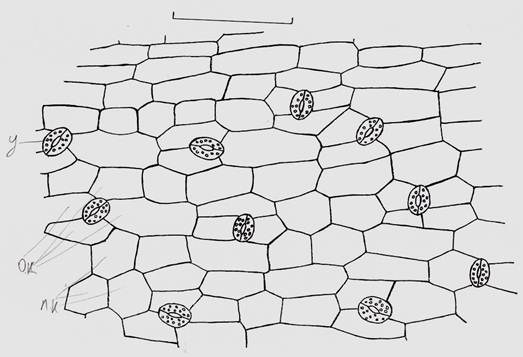
Рисунок 4. Анатомическое строение эпидермы листа: у. — устьица, о.к. — околоустьичные и п.к. — покровные клетки
Таким образом, рассматриваемые, завершающие молодые годичные побеги, верхушечные и пазушные «междоузлия» по морфологии и анатомии представляют собой удлиненные вальковатые листья с коронарно-центрическим типом хлоренхимы. Подобные конечные «междоузлия» заканчиваются тупозамкнутой верхушкой, а не точкой роста или почкой. Два слоя палисадной хлоренхимы и хлоренхимная кранц-обкладка образуют сплошное кольцо под эпидермой. К кранц-обкладке подходят мелкие почти чисто флоэмные проводящие пучки. В центре вальковатого листа проходят три наиболее крупных (по сравнению с периферическими) коллатеральных проводящих пучка. К основанию вальковатых листьев в центральных проводящих пучках усиливаются арматурные свойства: за счет склеренхимных тканей луба и древесины. Ориентацию в пространстве вальковатого листа до 27 мм длиной и 1,7 мм в диаметре обеспечивает не столько слабо развитая арматурная ткань, сколько тургор водозапасающих клеток. Устьица на эпидерме аномоцитного типа, до 125 штук на 1 мм2.
Таблица 1.
Морфометрического параметры клеток и тканей вальковатого листа C . aphyllym (май 2013 г)
|
Параметры листа (дл. * шир., мм) |
20—27*1,5 |
|
Высота эпидермы (мкм) |
14—20 |
|
Толщина кутикулы (мкм) |
4—5 |
|
Параметры покровных клеток эпидермы (дл. * шир., мкм) |
48*25 |
|
Параметры устьиц (мкм) |
26*16 |
|
Число устьиц (на 1мм2) |
125 |
|
Число околоустьичных клеток |
4(3) |
|
Параметры палисадных клеток: наружного слоя (мкм) |
27—30*10—14 |
|
Параметры палисадных клеток: внутренего слоя (мкм) |
30—32*10—14 |
|
Параметры клеток кранц-обкладки (дл. * шир., мкм) |
18—96*14—20 |
|
Параметры периферических проводящих пучков (дл. * шир.) |
24—30*30—70 |
|
Параметры медианного пучка: луб. скл.-мягкий луб – ксилема (мкм) |
45—30—40 |
|
Параметры латерального пучка: луб. скл.-мягкий луб – ксилема (мкм) |
30—30—40 |
|
Параметры водозапасающих клеток |
65—50 |
Список литературы:
1.Абдуллаева А.Т. Морфология цветка и биология цветения некоторых видов p. Calligonum L. Автореферат дисс. … к.б.н. Ташкент. 2000. — 16 с.
2.Ашурметов О.А. др. Формирование покровов плода и семени у Calligonum yunceum. (Polygonaceae). Ботан. ж. — 2006. — Т. 91. — № 9. — С. 1378—1388.
3.Васильев А.С. и др. Ботаника: морфология и анатомия растений. М.: Просвещение. 1988. — 480 с.
4.Зволинский В.П. и др. Способ создания долговременных плантаций Кандыма голова медузы Calligonum Caput-medusae Schrenk. Патент на изобретение RUS 2368112 от 28. 07. 2008.
5.Лачко О.А., Клеев В.М., Самтонов В.Б. Способ создания пастбищных угодий на песках. Номер патента: 2053617. Страна: Россия. Год 1996.
6.Парлиев Ю.П. Биология кандыма и опыт и его возделывания в условиях пустыни Узбекистана. Автореферат дисс. …к.б.н./ Ташкент: ТГУ. 1964. — 16 с.
7.Растительный мир Земли. под ред. Ф. Фукарека. Т. 2., М.: Мир 1982. — 320 с.
8.Сосков Ю.Д. Род Calligonum L — джузгун (систематика, география, эволюция). Автореферат дисс. … д.б.н./Ленинград,/1988.
9.Ranjbarfordoel A., Vandamme P., Samson R. Some ecophysiological characteristics of arta (Callogonum comosum L. Herit) in response to drought stress. Forest science and Practice. — 2013. — T. 15. — № 2. — C. 114—120.
10.Tavakkoli S., Osaloo S.K., Maassoumi A.A. The phylogeny of Callidonum and Pteropyrum (Polygonaceae) Baseol on nuclear ribosomal dna its and chloroplast TRNL-F sequences. Yranian yournal of Biotechnology. — 2010. — T. 8. — № 1. — C. 7—15.
дипломов