
Статья опубликована в рамках: XIV Международной научно-практической конференции «Инновации в науке» (Россия, г. Новосибирск, 19 ноября 2012 г.)
Наука: Биология
Скачать книгу(-и): Сборник статей конференции, Сборник статей конференции часть II
- Условия публикаций
- Все статьи конференции
дипломов

АНАЛЬГЕТИЧЕСКИЕ ПОЛИПЕПТИДЫ КУНИТЦ-ТИПА АКТИНИИ HETERACTIS CRISPA
Гладких Ирина Николаевна
канд. хим. наук, науч. сотр., ТИБОХ ДВО РАН,
г. Владивосток
Е-mail:
Чаусова Виктория Евгеньевна
мл. науч. сотр., ТИБОХ ДВО РАН,
г. Владивосток
Е-mail: v.chausova@gmail.com
Табакмахер Валентин Михайлович
аспирант, ТИБОХ ДВО РАН,
г. Владивосток
Е-mail: tabval@yandex.ru
Зелепуга Елена Александровна
канд. физ.-мат. наук, науч. сотр., ТИБОХ ДВО РАН,
г. Владивосток
Е-mail: zel@piboc.dvo.ru
Ткачева Екатерина Сергеевна
мл. науч. сотр., ТИБОХ ДВО РАН,
г. Владивосток
Е-mail:
Синцова Оксана Владимировна
студент Дальневосточного Федерального Университета,
г. Владивосток
Е-mail: sintsova0@mail.ru
Монастырная Маргарита Михайловна
д-р хим. наук, вед. науч. сотр., ТИБОХ ДВО РАН,
г. Владивосток
Е-mail: rita1950@mail.ru
Козловская Эмма Павловна
д-р хим. наук, профессор, ТИБОХ ДВО РАН,
г. Владивосток
Е-mail: kozempa@mail.ru
ANALGESIC KUNITZ-TYPE POLYPEPTIDES FROM THE SEA ANEMONE HETERACTIS CRISPA
Irina Gladkikh
PhD, researcher PIBOC FEB RAS,
Vladivostok
Victoria Chausova
Junior researcher PIBOC FEB RAS,
Vladivostok
Valentin Tabakmakher
PhD-student PIBOC FEB RAS,
Vladivostok
Elena Zelepuga
PhD, researcher PIBOC FEB RAS,
Vladivostok
Ekaterina Tkacheva
Junior researcher PIBOC FEB RAS,
Vladivostok
Oksana Sintsova
Studen of Far Eastern Federal University,
Vladivostok
Margarita Monastyrnaya
Doctor of Chemistry, Leading Researcher PIBOC FEB RAS,
Vladivostok
Emma Kozlovskaya
Doctor of Chemistry, Professor PIBOC FEB RAS,
Vladivostok
Работа выполнена при финансовой поддержке гранта РФФИ 12-08-31567 мол_а.
АННОТАЦИЯ
Для получения новых анальгетических полипептидов с помощью методов компьютерного моделирования из комбинаторной библиотеки полипептидов Кунитц-типа актинии Heteractis crispa выбраны две последовательности, HCGS 1.10 и 1.36. Показано, что единичные аминокислотные замены в полипептидах не препятствуют взаимодействию с TRPV1-рецептором. Методами молекулярного клонирования на основе вектора pET32b(+) созданы две генно-инженерные конструкции, несущие гены анальгетических полипептидов. Проведена аналитическая экспрессия гибридных белков. Показано, что полипептиды экспрессируются в растворимом виде.
ABSTRACT
To obtain new analgesic polypeptides using computer modeling methods two sequences HCGS 1.10 and 1.36 from a combinatorial library of Kunitz-type polypeptides of the sea anemone Heteractis crispa were selected. It was shown that point amino acid substitutions in the polypeptides do not prevent to the interaction with TRPV1-receptor. Using molecular cloning techniques based on vector pET32b (+) two constructs with the analgesic polypeptides genes were created. The analytical expression of the fusion proteins was performed. It was shown that the polypeptides are expressed in a soluble form.
Ключевые слова: комбинаторная библиотека; анальгетические полипептиды; компьютерное моделирование; молекулярное клонирование, экспрессия.
Keywords: combinatorial library; analgesic polypeptides; computer modeling; molecular cloning; expression.
Полипептиды Кунитц-типа широко представлены как в наземных, так и морских ядовитых организмах и являются компонентами не только ядов, но и секрета кожи. Они проявляют разнообразные виды биологической активности, действуют на различные биологические мишени (ферменты, ионные каналы и ионотропные рецепторы), могут играть важную роль в выживании животных, защите от деградации полипептидных токсинов и других белковых компонентов яда и/или в формировании синергетического эффекта действия компонентов яда. Согласно литературным данным ряд полипептидных молекул с разнообразными функциями, продуцируемых ядовитыми организмами, имеет Кунитц-фолд. Природа, вероятно, «отбирала» этот мотив как общий каркас для проявления различных видов биологической активности [10, 11, 15]. Установлено, что в ядах змей, скорпионов, пауков, конусов и актиний полипептиды Кунитц-типа продуцируются в виде многочисленных изоформ и образуют природные комбинаторные библиотеки. В настоящее время полипептиды Кунитц-типа представляют фундаментальный интерес, заключающийся в получении на их основе высокоспецифичных инструментов для исследования различных биологических мишеней и создания лекарственных препаратов нового поколения. Однако близкие значения физико-химических характеристик нативных полипептидов создают серьезную проблему при выделении данных соединений в индивидуальном состоянии. Кроме того, поиск и установление первичной структуры минорных изоформ полипептидов сложно осуществить методами белковой химии, которые требуют значительного количества исходного сырья.
В то же время анализ аминокислотных последовательностей и пространственных структур полипептидов комбинаторных библиотек с помощью методов молекулярной биологии и компьютерного моделирования позволяет быстро получить полную информацию о многообразии существующих изоформ и специфичности действия на биологические мишени. Эти современные методологические подходы дают возможность получения отдельных полипептидов, потенциальных кандидатов для создания фармакологических препаратов с заданными свойствами в рекомбинантной форме, что способствует сохранению численности популяций морских гидробионтов.
Ранее установлено, что 33 представителя HCGS-полипептидов образуют комбинаторную библиотеку полипептидов Кунитц-типа актинии Heteractis crispa [12]. С целью структурно-функционального анализа полученных структур для каждого GS-полипептида комбинаторной библиотеки с помощью программы SPDBV [16] был рассчитан молекулярный электростатический потенциал (МЭП) и построены эквипотенциальные поверхности, представляющие собой трехмерную визуализацию МЭП (рис. 1). МЭП является важной характеристикой белковой молекулы и определяет ее специфичность и кинетику молекулярного связывания [17].
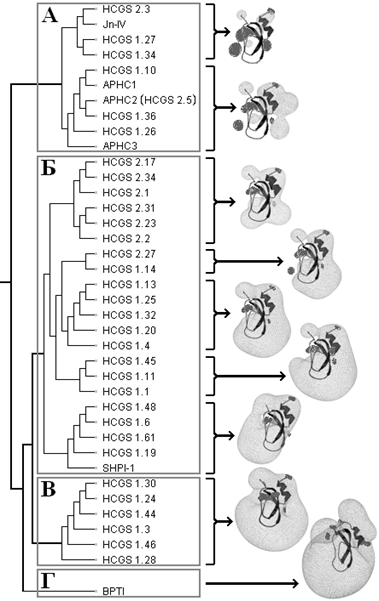
Рисунок 1. Дендрограмма полипептидов Кунитц-типа актинии H. crispa, SHPI-1 актинии Stichodactyla helianthus [7] и BPTI — полученная с применением программы PIPSA и сервера webPIPSA [17, 9]. Структуры макромолекул полипептидов представлены в виде ленточной диаграммы (обозначены черным цветом). Справа от дендрограммы представлены эквипотенциальные поверхности, характерные для данной группы полипептидов в кластерах А — Г. Расчеты МЭП выполнены в программе SPDBV (цветовое обозначение: темно-серый — отрицательный потенциал, светло-серый — положительный потенциал).
Как правило, полипептиды с близкими электростатическими свойствами имеют сходный механизм связывания с мишенями и, возможно, одинаковую биологическую активность. Было отмечено, что для HCGS-полипептидов характерно несколько типов МЭП. В результате проведенного анализа подобия МЭП представителей комбинаторной библиотеки, ингибитора протеиназ SHPI-1 из актинии Stichodactyla helianthus и BPTI — как наиболее изученного представителя структурного семейства Кунитца, полипептиды были разделены на четыре кластера (рис. 1).
Особый интерес представляют представители комбинаторной библиотеки, которые характеризуются точечным распределением МЭП и формируют кластер А. Идентичность последовательностей внутри данной группы составляет 84—98 %. В отличие от других представителей комбинаторной библиотеки HCGS, эти полипептиды имеют остатки Thr (положение 14) в центре реактивного сайта и Glu (положение 38) в сайте слабых взаимодействий с сериновыми протеиназами. В этом кластере можно выделить две подгруппы соединений: одна из них образована полипептидами, гомологичными ингибитору трипсина Jn-IV [3] со степенью идентичности аминокислотной последовательности внутри подгруппы 88—98 %, и другая — гомологичными анальгетическим полипептидам APHC1-APHC3 [5, 4] со степенью идентичности 91—98 %.
Принимая во внимание столь высокую степень идентичности аминокислотных последовательностей и общий характер распределения МЭП, мы можем предположить, что новые HCGS-полипептиды (1.10, 1.26, и 1.36), которые группируются с APHC1-APHC3 (рис. 1), должны, помимо трипсинингибирующей активности (Ki ~ 10-6—10-7 М), обладать способностью модулировать in vitro болевой ваниллоидный рецептор TRPV1 и оказывать анальгетическое действие in vivo. Поэтому для создания генно-инженерных конструкций были выбраны две последовательности GS-полипептидов, а именно HCGS 1.10 и HCGS 1.36, которые содержат положительно заряженные остатки Arg18 и Arg48 (рис. 2), вносящие, согласно литературным данным [5], существенный вклад в модулирование функциональной активности TRPV1-рецептора.
Jn-IV ----GSICLEPKVVGPCTAYFPRFYFDSETGKCTPFIYGGCEGNSYVDEKLHACRAICRA-----
HCGS 2.3 ----GSICLEPKVVGPCTAYFPGFYFDSETGKCTPFIYGRCEGNGNNFETLHACRAICRA-----
HCGS 1.27 ---—GSICLEPKVVGPCTAYFPRFYFDSETGKCTPFIYGGCEGNGNNFETLHACRAICRA-----
HCPG 1.34 ----GSICLEPKVVGPCTAYFPRFYFDSETGKCTPFIYGGCEGSGNNFETLHACRAICRA-----
HCGS 1.10 ----GSICLEPKVVGPCTAYLRRFYFDSETGKCTPFIYGGCEGNGNNFETLRACRAICRA-----
APHC1 ----GSICLEPKVVGPCTAYFRRFYFDSETGKCTVFIYGGCEGNGNNFETLRACRAICRA-----
HCGS 1.36 ----GSICLEPKVVGPCTAYFRRFYYDSETGKCTPFIHGGCEGNGNNFETLRACRAICRA-----
APHC2 ----GSICLEPKVVGPCTAYFRRFYFDSETGKCTPFIYGGCEGNGNNFETLRACRAICRA-----
APHC3 ----GSICLEPKVVGPCTAYFPRFYFNSETGKCTPFIYGGCEGNGNNFETLRACRGICRA-----
HCGS 1.26 ----GSICLEPKVVGPCTAYFRRFYFDSETGKCTPFIYGGCEGNGNNFETLHACRAICRA-----
HCGS 2.5 ----GSICLEPKVVGPCTAYFRRFYFDSETGKCTPFIYGGCEGNGNNFETLRACRAICRA-----
Рисунок 2. Множественное выравнивание аминокислотных последовательностей GS-полипептидов кластера А. Подчеркнуты полипептиды, входящие с нативными анальгетиками APHC1 и APHC2 [5, 4] в состав двух «анальгетических» подгрупп.
Механизм взаимодействия полипептидов HCGS 1.10 и HCGS 1.36 с TRPV1 рецептором был установлен на основании построенной ранее c помощью метода молекулярного докинга структурной модели комплекса APHC1-TRPV1 [1], а также проведенного in silico-мутагенеза APHC1 в комплексе с болевым рецептором. Согласно результатам молекулярного моделирования и молекулярной динамики, проведенным с использованием программы МОЕ, единичные негомологичные аминокислотные замены не должны оказывать существенного влияния на прочность связывания полипептидов данной группы с болевым рецептором.
Так полипептид HCGS 1.10 отличается от APHC1 несколькими заменами. Замена Phe17 на Leu локализована в участке ароматического кластера, который отличает группу полипептидов, обладающих анальгетической активностью, от других представителей комбинаторной библиотеки H. crispa полипептидов Кунитц-типа. В то время как замена Val31 на Pro также присутствует в полипептиде APHC3 и не приводит к утрате анальгетического действия полипептида. Молекулярно-динамические расчеты комплекса HCGS 1.10-TRPV1 в водном окружении при рН 6,0 показали, что замена Phe17Leu не привела к заметному изменению количественных характеристик взаимодействия с рецептором по сравнению с комплексом APHC1-TRPV1. При этом замена ароматического аминокислотного остатка Phe на алифатический Leu не препятствовала образованию кластера π- катионных взаимодействий между полипептидом и рецептором: Arg18-B:Phe721-B:Met717 и Ile 33-Tyr16-B:Arg718 (рис. 3).
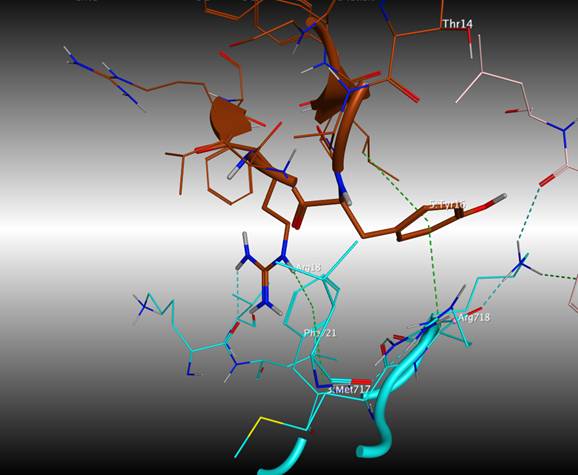
Рисунок 3. Межмолекулярные взаимодействия полипептида HCGS 1.10 с TRPV1. Аминокислотные остатки в комплексе представлены в виде стержневой диаграммы. Водородные связи и π-катионные взаимодействия показаны пунктирными линиями.
Ранее нами было показано, что Val31 анальгетического полипептида APHC1 образует водородную связь с Lys719 субъединицы D рецептора TRPV1 [1]. Однако присутствие остатка Pro в этом положении у всех полипептидов «анальгетической группы», за исключением APHC1, не создает пространственных или иных препятствий для взаимодействия с болевым рецептором. Это хорошо согласуется с экспериментальными данными об анальгетическом действии APHC3 [4].
Полипептид HCGS 1.36, помимо выше упомянутой замены в положении 31, содержит еще две замены: Phe22Tyr и Tyr34His. Несмотря на то, что эти замены отличают данный полипептид от всех остальных представителей группы, нами не выявлено сколь-нибудь важных изменений во взаимодействиях TRPV1 рецептора с полипептидом по сравнению с другими представителями группы. Phe22 полипептида HCGS 1.36 в составе комплекса с TRPV1 локализован на периферийной части области контактов, и его боковая часть не экспонирована в интерфейс межмолекулярных взаимодействий. По-видимому, влияние этого остатка может быть существенным лишь с точки зрения стабильности самого полипептида, но не его комплекса с TRPV1 рецептором. His34, так же как и Tyr34 в комплексе APHC1-TRPV1, не вовлечен во взаимодействие с рецептором, хотя его боковая цепь экспонирована в интерфейс в области Lys719 и Ala720 субъединицы B рецептора.
Таким образом, показано, что единичные аминокислотные замены в полипептидах HCGS 1.36 и HCGS 1.10 не представляют стерических или электростатических препятствий для взаимодействия с болевым рецептором TRPV1 и, следовательно, для проявления ими анальгетического действия in vivo. Поэтому данные полипептиды были отобраны для дальнейшего молекулярного клонирования с целью получения рекомбинантных форм и дальнейших исследований.
Широко распространенным способом получения белковых молекул является их функциональная экспрессия в клетках прокариот, например в Escherichia coli [18]. В условиях бактериальной системы высокий уровень продукции рекомбинантных полипептидов может достигаться за счет правильного выбора экспрессионной системы, оптимизации условий биосинтеза и очистки целевого белка. Наличие в рекомбинантных белках дисульфидных связей значительно осложняет продукцию функционально-активных белков, т.к. у E. coli отсутствуют ферментативные системы, отвечающие за образование дисульфидных связей и поэтому молекула такого полипептида, синтезированного в бактериальной клетке, не стабилизированная дисульфидными связями, становится очень чувствительной к протеолитической деградации [8]. Поэтому функциональную экспрессию дисульфид-содержащих белков, как правило, проводят в составе слитных конструкций в паре с геном белка-носителя тиоредоксина (Trx), который способен не только катализировать замыкание дисульфидных связей в целевом белке, но и удерживать его в растворимом состоянии, так как Trx накапливается в цитоплазме E. coli (до 40 % от общей массы клеточных белков), оставаясь при этом в растворимой форме. Кроме того, благодаря своим небольшим размерам (11675 Да) Trx, включенный в гибридный белок, не увеличивает чрезмерно его массу. Расположение на поверхности белковой молекулы Trx N- и C-концов удобно для присоединения к нему других белков [19, 13].
В линкере, соединяющем последовательности белка-носителя и целевого белка, как правило, предусмотрен сайт для последующего химического или ферментативного гидролиза гибридного белка. Гидролиз с помощью химического расщепления гибридного белка бромцианом (BrCN) позволяет получить целевой продукт с N-концевой последовательностью нативного полипептида [6]. Однако использование BrCN подходит только для белков, не содержащих собственных остатков метионина.
При выборе системы экспрессии было отдано предпочтение вектору pET32b(+), который используется для экспрессии в E. coli гибридных белков с Trx. Преимущество данной экспрессионной конструкции заключается в том, что вектор обеспечивает высокий выход целевого белка, а Trx — правильность замыкания дисульфидных связей, необходимых для формирования биологически активной структуры дисульфид-содержащих белков, к которым относятся полипептиды Кунитц-типа.
Нуклеотидные последовательности, кодирующие HCGS 1.10 и HCGS 1.36 были проверены на присутствие редких кодонов в клетке хозяина E. coli. Выбранные последовательности HCGS-полипептидов, уже содержащие на 5`-конце сайт для EcoRI рестриктазы, кодон метионина (Met), а на 3`-конце — стоп кодон (TAA) и сайт для XhoI рестриктазы, были субклонированы из вектора pTZ57R/T по сайтам рестрикции EcoRI и XhoI в вектор pET32b(+) в составе T7-контролируемого оперона (рис. 4).
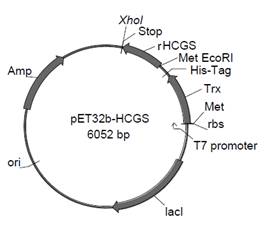
Риcунок 4. Физическая карта плазмиды pET-32b-HCGS полипептид.
Остаток Met был введен перед последовательностью целевого продукта, чтобы в дальнейшем эффективно провести гидролиз BrCN гибридного белка и отщепить зрелый полипептид с нативной N-концевой последовательностью от конечного слитного белка-носителя. Собственного остатка метионина аминокислотные последовательности HCGS-полипептидов не имеют, поэтому выбранный метод химического расщепления пептидной связи можно использовать без каких-либо ограничения. Фрагменты HCGS-полипептидов встраивали после фрагмента Trx и His6-Tag, аффинно-хелатирующего Ni2+-содержащую смолу, что позволяет на стадии металлоаффиной хроматографии избавиться от большинства балластных белковых компонентов клеточного лизата [3].
Далее полученными конструкциями, обозначенными как pET32-HCGS 1.10 и pET32-HCGS 1.36, трансформировали штамм E. coli Top 10 с помощью метода электропорации, который затем высевали на чашки c твердой питательной средой, содержащей селективный антибиотик ампициллин (100 мкг/мл). Положительные клоны, содержащие рекомбинантные плазмиды, были отобраны с помощью метода ПЦР «на колониях» и проверены на отсутствие мутаций прямым секвенированием с использованием стандартных праймеров. Затем результирующими генетическими конструкциями трансформировали штамм E. coli BL21(DE3) методом электропорации для проведения аналитической экспрессии. Аналитическую экспрессию рекомбинантных полипептидов проводили в 25 мл питательной культуры при интенсивной аэрации (180 об/мин) и температуре 37°С. Наработку целевого слитного белка индуцировали добавлением 1 мМ ИПТГ. Электрофоретическое разделение компонентов клеточных лизатов в ДСН-ПААГ показало, что молекулярные массы рекомбинантных гибридных полипептидов соответствовали расчетным данным (больше 26 кДа, рис. 5). Анализ растворимых и нерастворимых фракций бактериальных лизатов показал, что рекомбинантные гибридные HCGS-полипептиды содержатся только в растворимых фракциях.
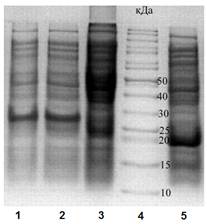
Рисунок 5. Электрофореграмма гибридных белков. Клеточные лизаты BL21(DE3) штаммов-продуцентов E. coli: 1 — pET32b- rHCGS1.10; 2 — pET32b-rHCGS1.36; 3 — контроль (pET32b-Trx без добавления ИПТГ); 4 — белки-стандарты молекулярной массы; 5 — контроль pET32b-Trx.
Таким образом, в результате проделанной работы созданы две генно-инженерные конструкции, несущие гены анальгетических HCGS-полипептидов, на основе вектора pET32b(+) и проведена аналитическая экспрессия гибридных рекомбинантных белков Кунитц-типа актинии H. crispa. Показано, что два гибридных рекомбинантных HCGS-полипептида экспрессируются в растворимом виде.
Список литературы:
1.Зелепуга Е.А., Табакмахер В.М., Чаусова В.Е., Монастырная М.М., Исаева М.П., Козловская Э.П. Взаимодействие полипептидов кунитц-типа актинии Heteractis crispa с болевым ваниллоидным рецептором ТRPV1: in silico исследование // Биоорг. химия. — 2012. — Т. 38, C. 185—198.
2.Зыкова T.A., Винокуров Л.M., Mаркова Л.Ф., Koзловская Э.П., Еляков Г.Б. Аминокислотная последовательность ингибитора трипсина IV из Radianthus macrodactylus // Биоорг. xимия. — 1985. — Т. 11, С. 293—301.
3.Иванов А.С., Згода А.И., Арчаков В.Г. Технологии белковой интерактомики // Биоорг. химия. — 2011. — Т. 37, № 1. С. 8—21.
4.Козлов С.А., Андреев Я.А., Мурашев А.Н., Скобцов Д.И., Дьяченко И.А., Гришин Е.В. Новые полипептидные компоненты с анальгетической активностью из морской анемоны Heteractis crispa // Биоорг. химия. — 2009. — Т. 35. С. 789—798.
5.Andreev Y.A., Kozlov S.A., Koshelev S.G., Ivanova E.A., Monastyrnaya M.M., Kozlovskaya E.P., Grishin E.V. Analgesic compound from sea anemone Heteractis crispa is the first polypeptide inhibitor of vanilloid receptor 1 (TRPV1) // J. Biol. Chem.— 2008. — V. 283. P. 23914—23921.
6.Andreev Y.A., Kozlov S.A., Vassilevski A.A., Grishin E.V. Cyanogen bromide cleavage of proteins in salt and buffer solutions // Anal Biochem. — 2010. — V. 407, N 1. P. 144—146.
7.Antuch W., Berndt K.D., Chavez M.A., Delfín J., Wüthrich K. The NMR solution structure of a Kunitz-type proteinase inhibitor from the sea anemone Stichodactyla helianthus // Eur. J. Biochem. — 1993. — V. 212, P. 675—684.
8.Berndt C., Lillig C.H., Holmgren A. Thioredoxins and glutaredoxins as facilitators of protein folding // Biochim. Biophys. Acta. — 2008. — V. 1783, N 4. P. 641—650.
9.De Rienzo F., Gabdoulline R.R., Menziani M.C., De Benedetti P.G., Wade R.C. Electrostatic Analysis and Brownian Dynamics Simulation of the Association of Plastocyanin and Cytochrome F // Biophys. J. — 2001. — V. 81, P. 3090—3104.
10.Escoubas P. Molecular diversification in spider venoms: a web of combinatorial peptide libraries // Mol. Divers. — 2006. — V. 10. P. 545—554.
11.Fry B.G. From genome to «venom»: molecular origin and evolution of the snake venom proteome inferred from phylogenetic analysis of toxin sequences and related body proteins // Genome Res. — 2005. — V. 15, N 3. P. 403—420.
12.Isaeva M.P., Chausova V.E., Zelepuga E.A., Guzev K.V., Tabakmakher V.M., Monastyrnaya M.M., Kozlovskaya E.P. A new multigene superfamily of Kunitz-type protease inhibitors from sea anemone Heteractis crispa // Peptides. — 2012. — V. 34. P. 88 — 97.
13.McCoy J., Lavallie E. Expression and purification of thioredoxin fusion proteins // Curr. Protoc. Mol. Biol. — 2001. — Chapter 16:Unit16.8.
14.Novagen. pET System Manual. 2008. 11th edition.
15.Olivera B.M., Hillyard D.R., Marsh M., Yoshikami D. Combinatorial peptide libraries in drug design: lessons from venomous cone snails // Trends Biotechnol. — 1995. — V. 13, N 10. P. 422—426.
16.Peitsch M.C. Protein modeling by E-mail // Nat. Biotechnol. 1995. V. 13. P. 658—660.
17.Richter S., Wenzel A., Stein M., Gabdoulline R.R., Wade R.C. webPIPSA: a web server for the comparison of protein interaction properties // Nucleic Acid Res. — 2008. — V. 36, P. 276—280.
18.Rudolph R. Successful protein folding on an industrial scale // Protein engineering: principles and practice. New York: Wiley-Liss. — 1996. P. 283—298.
19.Yasukawa T., Kanei-Ishii C., Maekawa T., Fujimoto J., Yamamoto T., Ishii S. Increase of solubility of foreign proteins in Escherichia coli by coproduction of the bacterial thioredoxin // J. Biol. Chem. — 1995. — V. 270, N 43. P. 25328—25331.
дипломов