
Статья опубликована в рамках: VII Международной научно-практической конференции «Научное сообщество студентов XXI столетия. ЕСТЕСТВЕННЫЕ НАУКИ» (Россия, г. Новосибирск, 07 февраля 2013 г.)
Наука: Биология
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
дипломов

СИНАПТИЧЕСКИЕ ОТВЕТЫ, ВЫЗЫВАЕМЫЕ СТИМУЛЯЦИЕЙ ЛЕМНИСКАЛЬНЫХ СИНАПСОВ В РЕЛЕЙНОМ ТАЛАМУСЕ НОВОРОЖДЕННЫХ МЫШЕЙ IN VITRO
Королева Ксения Сергеевна
студент 5 курса, кафедры физиологии человека и животных, Институт фундаментальной медицины и биологии К(П)ФУ, г. Казань
E-mail: kseniya.ks29061991@yandex.ru
Яковлев Алексей Валериевич
научный руководитель, канд. биолог. наук, доцент К(П)ФУ, г. Казань
Важной особенностью ранних этапов развития ЦНС является генетически обусловленная избыточность в образовании количества нейронов, их отростков и межнейронных контактов. По мере созревания ЦНС избыточность постепенно устраняется. Гибель (выборочная элиминация) лишних нейронов, так называемый апоптоз, служит устранению избыточных отростков и синапсов и выступает как один из способов «уточнения» плана формирования нервной системы [1, с. 271, 2, с. 121].
Эти процессы хорошо изучены на примере ядер таламуса, где происходит переключение специфической и неспецифической сенсорной информации от периферических рецепторов в кору головного мозга. Например, показано, что синаптические связи между ганглиозными клетками сетчатки и нейронами латеральных коленчатых тел таламуса в течение постнатального развития подвергаются обширной реорганизации [5, с. 603, 6, с. 29]. До открытия глаз, каждый нейрон коленчатых тел таламуса мыши получает до 20 синаптических входа от ганглиозных клеток сетчатки. Но уже через 2 недели после открытия глаз, происходит уменьшение количества синапсов до 1—3 [3, с. 956].
Аналогичные стадии процесса элиминации выявлены и в других сенсорных ядрах таламуса. Показано, что на второй неделе после рождения у грызунов происходит постепенно уменьшение количества синапсов и их созревание [2, с. 122]. Однако о процессах, происходящих в первую неделю постнатального развития данных не достаточно. В связи с этим, целью данной работы является оценка состава афферентных (сенсорных) входов в таламическое представительство вибрисс мыши в течение первой постнатальной недели.
Объект и методы исследования.
Эксперименты проводились на горизонтальных срезах головного мозга мышей, обоих полов в возрасте Р1 — Р7 (день рождения Р0), с соблюдением всех биоэтических норм. Мышей декапитировали, после чего, мозг быстро извлекался из черепной коробки. Сразу после извлечения, мозг охлаждался в растворе искусственной спинномозговой жидкости (artificial cerebral spinal fluid — ACSF), насыщенный кислородом (95 %) и углекислым газом (5 %). Горизонтальные срезы, толщиной 400 µм были получены на микротоме (Thermo Scientic Microm HM 650 V). Перед началом записи, срезы инкубировались в растворе ACSF, при комнатной температуре (21—23ºС) в течение часа. Все эксперименты выполнены в условиях наружной перфузии препарата. ACSF содержал в мМ: 126 NaCl, 3,5 KCl, 1,2 NaH2PO4*H2O, 1,3 MgCl2*6H2O, 2 CaCl2*6H2O, 26 NaHCO3, 10 глюкоза, насыщался 95 % О2 и 5 % СО2 .
В эксперименте использовали следующие вещества: D-(-) амино-5-фосфонопентаноат (d-APV, 40 µМ), 6 -циано-7 нитрокиноксалин (CNQX, 10 µМ), бикукулин (10 µМ) и габазин (10 µМ) (Sigma, США). Во время эксперимента вещества добавляли в перфузию в стандартный раствор ACSF.
Перед началом регистрации срезы просматривались с использованием 4х-объектива микроскопа Leica MZ6 (Германия). Для регистрации выбирались срезы, содержащие медиальный лемниск (ML) и вентральное базальное ядро (VPM) таламуса.
Под визуальным контролем стимулирующий биполярный металлический электрод размещался в ML. Для регистрации возбуждающих постсинаптических токов (ВПСТ) использовали стандартную микроэлектродную технику фиксации потенциала на целой клетке (whole-cell) при помощи усилителя Axopatch 200A (Axon Instruments) и программы AxoScope 9.2. Регистрация ВПСТ осуществлялась с помощью стеклянного микроэлектрода, заполненного пипеточным раствором, следующего состава (мМ): 135 Cs-gluconate, 2MgCl2,0,1 CaCl2, 1 EGTA, 10 HEPES, pH 7.25. Кроме того, пипеточный раствор содержал блокатор Na-каналов — производное лидокаина, QX-314. Сопротивление электрода составляло от 5 до 8 МΩ. Для стимуляции ML использовался импульсный генератор Master — 8 (A.M.P.I., Jerusalem, Israel). Стимул подавался с частотой 0,5 Гц. Для усреднения и обработки данных использовались программы Clampfit и Origin 7,5.
Результаты и обсуждение.
Во время стимуляции лемнискального пути, происходит высвобождение глутамата, вызывающего возбуждающие глутаматные токи. При фиксации мембранного потенциала клетки на уровне — 70 мВ, пороговая стимуляция одиночного волокна вызывала моносинаптический ответ с быстрой фазой роста и спада, который ингибировался специфическим блокатором АМПА-рецепторов — CNQX. При стимуляции с амплитудой 30 мкА происходило уменьшение АМПА-компонента ВПСТ с — 120.81±9.6 рА до —4.05±2.7 рА (n=5, p<0.05) после аппликации ингибитора, соответственно. На уровне мембранного потенциала + 40 мВ наблюдался синаптический ответ с медленной фазой спада, которая исчезала при аппликации ингибитора НМДА-рецепторов — d-APV. Так аппликация блокатора приводило к снижению амплитуды ВПСТП с 175.51±3.68 рА, до 3.35±1.72 рА (n=5, p<0.05).
Таким образом, в релейных нейронах VPM таламуса при стимуляции лемнискального пути были зарегистрированы ВПСТ, состоящие из АМПА- и НМДА-компонентов.
Для того чтобы определить действительно ли регистрируемые ВПСТ возникают в ответ на стимуляцию ML, проводился высокочастотный тест. При стимуляции лемнискального пути пачкой импульсов, состоящей из 5 сигналов с частотой следования 50 Гц, наблюдалось угнетение амплитуды вызванных ответов на каждый последующий стимул. Амплитуда ВПСТ на первый стимул составила 236.16±8.71 pA и на последний — 105.69±4.1 pA (n=5, p< 0.05). Таким образом, можно сделать вывод о том, что при стимуляции пачкой из 5 сигналов, происходит угнетение каждого последующего вызванного сигнала, что является критерием определения стимуляции лемнискального пути в наших экспериментах.
Для анализа процесса элиминации синапсов исследовали количество афферентных входов в VPM ядро таламуса. Регистрировались синаптические ответы во время градуального увеличения интенсивности стимула при фиксации мембранного потенциала на уровне — 70 мВ и + 40 мВ, что позволяло анализировать АМПА- и НМДА-компоненты ВПСТ. Оценивали число афферентных входов по количеству дискретных ступенек в синаптическом ответе. В случае, когда приращения амплитуды ВПСТ были небольшими и множественными, число афферентных входов рассчитывалось как отношение максимальной амплитуды синаптического ответа при фиксации мембранного потенциала на уровне к амплитуде сигнала, вызываемого пороговой силой стимула. Так в первую неделю постнатального развития повышение силы стимуляции приводило к ступенчатому увеличению амплитуды ВПСТ за счет активации соседних волокон лемнискального пути. На рисунке 1 показаны изменения амплитуды АМПА- и НМДА-компонентов синаптических токов в первый (рис. 1А) и пятый (рис. 1 Б) день постнатального развития животного. Видно, что на Р1 происходит постепенное увеличение амплитуды ВПСТ в ответ на увеличение интенсивности стимула со множеством дискретных шагов. На Р5 также наблюдалось ступенчатое увеличение амплитуда АМПА- и НМДА-компонентов синаптических токов нейронов VPM, но с меньшим количеством дискретных ступенек в ответ на максимальную интенсивность стимула раздражения.
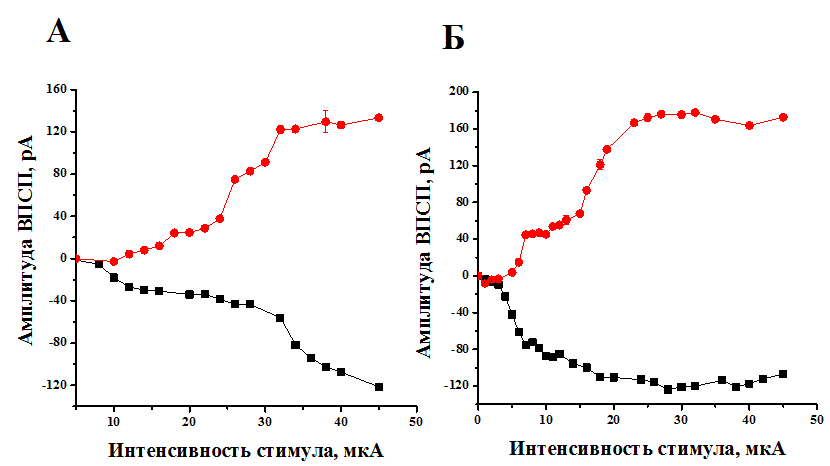
Рисунок 1. Изменение амплитуды АМПА- и НМДА-компонентов ВПСТ в зависимости от интенсивности стимула на первый (А) и пятый (Б) постнатального развития животного. (●) АМПА-токи ВПСТ, (■) НМДА-токи ВПСТ
Расчет количества сенсорных входов показал, что один нейрон VPM ядра таламуса в возрасте P1 иннервируется 5—12 волокнами медиального лемнискального пути, а к концу первой недели постнатального периода наблюдается уменьшение числа волокон до 4—6.
Таким образом, в первую неделю постнатального развития происходит уменьшение количества лемнискальных аксонов и увеличение вклада каждого аксона в синаптический ответ, имеющее большое значение для формирования карты тела в соматосенсорной коре.
Список литературы:
1.Равич-Щербо И.В. Психогенетика: учебное пособие для студентов высших учебных заведений, обучающихся по специальности и направлению «Психология». М.: издательство «Аспект Пресс», 2000. — 271 с.
2.Arsenault D., Zhang Z. Developmental remodelling of the lemniscal synapse in the ventral basal thalamus of the mouse. 2006. — v. 573 — P. 121—132.
3.Chen C., Regehr WG. Developmental remodeling of the retinogeniculate synapse. Neuron, 2000. — v. 28 — P. 955—966.
4.Shatz C. J. Emergence of order in visual system development. Proc. Natl. Acad. Sci. USA, 1996. — v. 93 — P. 602—608.
5.Wong R. O. Retinal waves and visual system development. Annu Rev. Neurosci, 1999. — v. 22 — P. 29—47.
дипломов
Оставить комментарий