
Статья опубликована в рамках: IX Международной научно-практической конференции «Научное сообщество студентов XXI столетия. ЕСТЕСТВЕННЫЕ НАУКИ» (Россия, г. Новосибирск, 04 апреля 2013 г.)
Наука: Медицина
Скачать книгу(-и): Сборник статей конференции
- Условия публикаций
- Все статьи конференции
отправлен участнику

ЭПИДЕМИОЛОГИЯ И РОЛЬ ВИРУСА ЭПШТЕЙНА-БАРР В РАЗВИТИИ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ
Дудукина Елена Александровна
студент 5 курса медико-профилактического факультета, Первый МГМУ им. И.М. Сеченова, г. Москва
E-mail: evpatora@yahoo.com
Львов Николай Дмитриевич
научный руководитель, канд. мед. наук, профессор, ФГБУ «НИИ вирусологии им. Д.И. Ивановского» Минздрава России, г. Москва
Вирус Эпштейна-Барр (ВЭБ), будучи одним из древнейших, как полагает современная наука, коэволюционаровал одновременно со своими хозяевами на протяжении миллионов лет [1, p. 929]. Благодаря способности поддерживать латентную инфекцию в организме в течение всей его жизни и периодически реактивироваться, не вызывая выраженных клинических проявлений у большинства инфицированных лиц, Эпштейн-Барр вирусная инфекция стала практически повсеместно распространена в популяции. Инфицированность мирового населения составляет >90 % [1, p. 986].
Дети в развивающихся странах приобретают Эпштейн-Барр вирусную инфекцию на первом году жизни, средний возраст сероконверсии в мире — 3—4 года. В развитых странах инфицирование происходит в подростковом возрасте [1, p. 929]. В некоторых развитых странах наблюдается бимодальный подъем заболеваемости с первым пиком у детей до 5 лет и вторым — среди детей 10-ти лет. Клинические проявления различны в зависимости от возраста первичного контакта с инфекционным агентом. При заражении в раннем детстве клиника стертая либо отсутствует, в то время как при инфицировании Эпштейн-Барр вирусом в школьном и подростковом возрасте развивается классическая клиническая картина инфекционного мононуклеоза: фебрильная лихорадка, ангина, увеличение заднешейных лифатических узлов, гепатоспленомегалия, в крови обнаруживаются атипичные мононуклеары. При использовании серологического метода диагностики с момента развития клинических признаков болезни на протяжении следующих 4—6 недель обнаруживаются IgM к VCA (viral capsid antigen) — вирусному капсидному антигену, с первой недели болезни до нескольких лет после выздоровления — Anti-EBV-EA-D Ig G (early antigen — D EBV) — антитела к раннему антигену D Эпштейн-Барр вируса, через несколько недель после появления клинических признаков определяются EBNA-IgG (иммуноглобулины класса G к нуклеиновым кислотам Эпштейн-Барр вируса), сохраняющиеся на протяжении всей жизни человека.
Титры антивирусных антител у серопозитивных лиц варьируются в зависимости от возраста, следуя U-образной кривой. Максимальные титры наблюдаются у детей и в возрастной группе старше 50 лет. Высокий уровень антивирусных антител среди детей объясняется наличием первичной инфекции, тогда как у пожилых людей повышение титра антител в основном связано с возраст-зависимой реактивацией инфекции вследствие ослабленного клеточного иммунного ответа [1, p. 929].
Эпштейн-Барр вирусная инфекция была обнаружена среди всех народов мира. В популяции циркулируют два основных типа вирусов, отличающихся по набору генов, которые кодируют ядерные белки в период латентного течения инфекции [1, p. 475]. Встречаемость штамма EBV1(Эпштейн-Барр вируса 1-го типа) несколько десятилетий назад была выше среди Западных популяций, тогда как штамм EBV2 (Эпштейн-Барр вирус 2-го типа) был распространен среди населения Африки, Папуа-Новой Гвинеи, а также среди коренного населения Аляски [1, p. 475]. Микст-инфекция двумя штаммами Эпштейн-Барр вируса отмечалась у ВИЧ/СПИД-положительных лиц. Однако информация, представленная в современных исследованиях, говорит о том, что большая часть населения всех континентов в настоящий момент подвержена инфицированию обоими типами Эпштейн-Барр вируса одновременно.
Распространенность полиморфизма специфических ДНК последовательностей Эпштейн-Барр вируса также имеет географические особенности. Наиболее изучен в этой области Эпштейн-Барр вирус-кодируемый онкоген LMP1 (latent membrane protein 1) — латентный мембранный белок 1 типа, различные варианты нуклеотидных последовательностей которого обнаруживаются у отдельных изолятов (штаммов) вируса, некоторые из них связаны с повышенным риском возникновения назофарингеальной карциномы [1, p. 929].
Недавние исследования показали, что вариации нуклеотидных последовательностей, кодирующих LMP1, полученные из клеток назофарингеальной карциномы в странах с высокой заболеваемостью этим новообразованием (в основном Юго-Восточная Азия), имеют четкие отличия от LMP1 белка клеток назофарингеальной карциномы, встречающейся в странах с низкой инцидентностью. Это дало основания предполагать существование определенной положительной селекции нуклеотидных последовательностей, кодирующих LMP1, в сторону сохранения изолятов с более выраженным онкогенным потенциалом в странах, эндемичных по назофарингеальной карциноме [1, p. 932].
Ранняя первичная Эпштейн-Барр вирусная инфекция связана, прежде всего, с плохими социально-экономическими условиям. Так в развитых странах подъем заболеваемости инфекционным мононуклеозом начинается гораздо позже, чем в развивающихся. Среди детей в возрасте от 5 до 10 лет, проживающих в неблагоприятных гигиенических условиях, серопревалентность по антивирусным антителам составляет 60 % по сравнению с менее чем 20 %-ой серопревалентностью среди детей, проживающих в более благоприятных санитарно-гигиенических условиях [1, p. 930].
Высокая распространенность Эпштейн-Барр вирусной инфекции наблюдается по всему миру, что служит индикатором весьма ограниченного влияния хозяин-специфических факторов на ее течение. Тем не менее, среди Инуитов (Эскимосов), проживающих в Антарктике, а также среди народов Юго-Восточной Азии существуют определенные генетические особенности иммунного ответа против Эпштейн-Барр вируса, что подтверждается необычно высоким титром антивирусных антител класса G к VCA (viral capsid antigen) — вирусному капсидному антигену, который остается постоянно высоким на протяжении всей жизни Инуитов [1, p. 931].
Эпштейн-Барр вирусная инфекция передается аэрозольным, фекально-оральным, контактным и трансмиссивным механизмами, реализуясь различными путями. Наиболее распространенным фактором передачи является слюна. Среди подростков и взрослого населения главным путем инфицирования является прямая передача вируса со слюной во время поцелуя, тогда как главным путем передачи инфекции среди маленьких детей является контактный, опосредованный через игрушки, на которых присутствует слюна носителя инфекции. В развивающихся странах были описаны случаи, когда матери перед тем, как давать пищу детям, пережевывали ее, что являлось главным фактором в распространении Эпштейн-Барр вирусной инфекции среди маленьких детей в семьях. Тем не менее, несмотря на 40-летнюю историю изучения Эпштейн-Барр вирусной инфекции, не все особенности передачи вируса до конца выяснены. После заражения инфицированное лицо может выделять Эпштейн-Барр вирус в окружающую среду различными путями. Так наличие Эпштейн-Барр вируса в секрете слюнных желез серопозитивных лиц варьируется от 22 % до 90 %. Наличие вирионов Эпштейн-Барр вируса в секрете слюнных желез, взятом в любой момент времени у конкретного пациента также может варьироваться в широких пределах от 32 % до 73 %. В ходе исследований не удалось обнаружить связь между наличием Эпштейн-Барр вируса в слюнном секрете и уровнем атипичных мононуклеаров (peripheral blood mononuclear cells, PBMCs). Т. о. полагают, что факторы ответственные за активацию репликации вируса и увеличение вирусной нагрузки в клетках слюнных желез слизистой оболочки ротоглотки и лимфоцитах крови могут различаться. Эпштейн-Барр вирус обнаруживается в цервикальном секрете 8—28 % девушек-подростков и женщин детородного возраста, а также в образцах спермы. Трансплацентарная передача Эпштейн-Барр вируса и передача с грудным молоком были описаны некоторыми исследователями, тем не менее, считается, что они не играют существенной эпидемиологической роли. Важными способами передачи Эпштейн-Барр вируса являются трансфузионный и трасплантационный. К примеру, эритроцитарная масса в среднем содержит два вириона Эпштейн-Барр вируса, в то время как в цельной крови среднее число вирионов колеблется от 600 до 700 ед. Эпштейн-Барр вирусная инфекция у людей после трансплантации органов в высшей степени коррелирует с возникновением так называемой посттрансплантационной лимфопролиферативной болезни (PTLD, posttransplant lymphoproliferative disease, or disorder), относящейся к опухолям [1, p. 931].
За последнее десятилетие методы, позволяющие обнаружить и количественно оценить внутри- и внеклеточную вирусную нагрузку, значительно усовершенствовались, что позволяет прослеживать в динамике вирусную нагрузку у пациентов, страдающих Эпштейн-Барр вирус-ассоциированными заболеваниями. Нагрузка более чем 102,5 копий Эпштейн-Барр вирусного генома /мкг ДНК при использовании метода ПЦР в режиме реального времени является клинически значимой [1, p. 932].
В крови клинически здоровых лиц с латентно протекающей Эпштейн-Барр вирусной инфекцией наблюдается приблизительно постоянное число инфицированных B-лимфоцитов, которое составляет 1—50 зараженных клеток на 1,000,000 B-лимфоцитов, вирусная нагрузка у таких лиц — менее 100 копий ДНК на 105 клеток. Однако даже у клинически здоровых лиц уровень вирусной нагрузки в периферической крови подвержен постоянным колебаниям. Как предполагают исследователи, это связано с эпизодической реактивацией вирус-зависимой клеточной пролиферации пораженных клеток: B-, Т-лимфоцитов, NK-клеток, макрофагов, нейтрофилов. В период острой первичной инфекции виремия достигает 103,7 копий генома ВЭБ /мкг ДНК, у пациентов с хронической активной формой Эпштейн-Барр вирусной инфекции, при назофарингеальной карциноме, посттрансплантационных лимфопролиферативных заболеваниях и других состояниях, связанных с персистенцией и последующей губительной реактивацией Эпштейн-Барр вируса, виремия — 104,1 копий /мкг ДНК [2]. Золотым стандартом в диагностике ВЭБ — ассоциированных заболеваний в настоящее время является обнаружение ДНК Эпштейн-Барр вируса в крови пациента с использованием ПЦР методики, дополнительное применение серологической диагностики проводят в случае необходимости [1, p. 932—934].
Многие из состояний, ассоциированных с Эпштейн-Барр вирусной инфекцией, развиваются у иммунокомпрометированных лиц, лиц пожилого возраста и в условиях, провоцирующих снижение иммунорезистентности организма. Для эпидемиологии многих Эпштейн-Барр вирус-положительных злокачественных новообразований характерна связь с географическим фактором — эндемичными районами, а также с генетическими детерминантами и влиянием окружающей среды. За последние десять лет были проведены исследования, в результате которых открылись новые подробности течения латентных состояний, закономерности опухолевой трансформации и механизмы иммортализации клеток. Латентное течение Эпштейн-Барр вирусной инфекции может иметь несколько форм, отличных по набору экспрессируемых генов, белковому профилю пораженных клеток, наличию или отсутствию характерных рецепторных белков на их поверхности, а также по морфологической картине пораженной ткани, что приводит к развитию различных ассоциированных состояний.
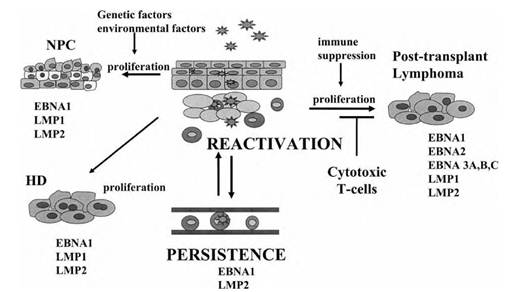
Рисунок 1. Патогенез ВЭБ инфекции
Классический путь заражения ВЭБ реализуется через слюну, при попадании на слизистую оболочку ротоглотки ВЭБ инвазирует эпителиоциты и лимфоциты, где размножается и реализует литический цикл. Вследствие виремии происходит диссеминация ВЭБ в красный костный мозг, органы РЭС. Также распространение ВЭБ инфекции происходит при миграции B-лимфоцитов. Реактивация вирусной инфекции возможна во всех инфицированных клетках, однако наибольшую роль во вторичном распространении ВЭБ играет механизм персистенции в B- лимфоцитах. На рисунке изображены паттерны экспрессии генов ВЭБ при различных формах течения латентной инфекции [1, p. 985].
Персистенция Эпштейн-Барр вируса в B-клетках памяти обычно протекает достаточно вяло с экспрессией лишь ядерного антигена 1 (nuclear antigen 1) — EBNA1 и/или латентного мембранного протеина 2 (latent membrane protein) — LMP2. Пролиферация пораженных B-лимфоцитов подконтрольна цитотоксическим лимфоцитам, распознающим EBNA белки как антигены. Именно это обстоятельство является объяснением высокой частоты развития B-лимфом у пациентов в состоянии иммунодепрессии, когда функция цитотоксических лимфоцитов подавлена и контроль за пролиферацией B-лимфоцитов утрачен. Реактивация литического цикла Эпштейн-Барр вируса в эпителиоцитах слизистой оболочки ротоглотки приводит к распространению инфекции в популяции, а латентное течение инфекции в эпителиоцитах ротоглотки с экспрессией генов EBNA1, LMP1 и LMP2 обуславливает синтез белков, трансформирующих и иммортализирующих клетки, приводя к возникновению различных типов назофарингеальной карциномы [1, p. 987].
В течение латентного периода Эпштейн-Барр вирусной инфекции происходит экспрессия многих генов ВЭБ. In vivo принято выделять три формы течения латентной инфекции в зависимости от набора экспрессируемых вирусных генов в пораженных клетках. В клетках лимфобластоидных культур in vitro (lymphoblastoid cell lines, LCL) и в клетках посттрансплантационных лимфом in vivo экспрессируются гены шести основных белков латентной инфекции: EBNA 1, 2, 3A, 3B, 3C, и ген лидирующего протеина — EBNA-LP (Leader Protein). Перечисленные белки регулируют экспрессию собственных генов, а также генов, кодирующих LMP1 и LMP2. Кроме того в клетках посттрансплантационных лимфом обнаруживаются неполиаденилированные РНК — EBER1 и EBER2. Они представляют собой молекулы некодирующей РНК длиной около 170 нуклеотидов. Транскрипция этих РНК молекул осуществляется РНК-полимеразой III клетки-хозяина. Биологическим действием EBERs является увеличение продукции ИЛ-10 Th2-лимфоцитами и угнетение функции цитотоксических T-лимфоцитов. Также в клетках LCL и посттрансплантационных лимфом обнаруживаются продукты правосторонней транскрипции BamHI A региона генома Эпштейн-Барр вируса — BARTs молекулы, которые представляют собой группу из 20 различных видов молекул микроРНК, получившихся в результате сплайсинга первичной молекулы РНК. BARTs активно участвуют в регуляции экспрессии различных вирусных (например, LMP1) и клеточных генов. Латентная инфекция, сопровождающаяся экспрессией всех перечисленных вирусных генов, считается латентной инфекцией 3 типа. Первые два типа течения латентной инфекции не отличаются подобным разнообразием экспрессируемых генов Эпштейн-Барр вируса в пораженных клетках. 1 тип течения латентной инфекции является наименее активным в отношении экспрессии вирусных генов, однако это не означает, что данный тип персистенции имеет наиболее благоприятное течение. Характерным признаком персистенции 1 типа является экспрессия генов EBNA1 и присутствие EBERs вследствие транскрипции BamHI A региона генома ВЭБ. Данный тип персистенции Эпштейн-Барр вируса встречается в клетках лимфомы Беркитта. 2 тип течения латентной Эпштейн-Барр вирусной инфекции был найден в малигнизированных клетках назофарингеальной карциномы и Ходжкинской лимфомы, при этом набор экспрессируемых генов ограничивается следующими: EBNA1, LMP1, LMP2, EBERs, BARTs [1, p. 987—983].
Таблица 1.
Тип течения латентной инфекции в зависимости от паттерна экспрессии генов Эпштейн-Барр вируса
|
Экспрес сируемый ген |
EBNA -1 |
EBNA -2A/B |
EBNA-3A EBNA-3B EBNA-3C |
EBNA -LP |
LMP -1 |
LMP -2A |
LMP -2B |
EBERs |
BARTs |
||
|
Продукт |
белок |
РНК |
микро РНК |
||||||||
|
Лат. 1 |
+ |
- |
- |
- |
- |
- |
- |
+ |
+ |
||
|
Лат. 2 |
+ |
- |
+ |
+ |
+ |
+ |
+ |
+ |
|||
|
Лат. 3 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
В случае опухолей, ассоциированных с ВЭБ-инфекцией, EBNA2 и EBNA3 экспрессируются в клетках при посттрансплантационных лимфопролиферативных заболеваниях. Благодаря экспрессии этих белков, являющихся мишенями для специфицеских цитотоксических лимфоцитов (CTL), возможно успешное лечение посттрансплантационных лимфом с помощью иммунотерапии. Таким образом, экспрессия протеинов LMP1 и 2 в отсутствие экспрессии EBNA2 и EBNA3, белков-мишеней для T-киллеров, является одним из факторов, способствующих развитию злокачественных опухолей. Резюмируя все сказанное выше, можно сделать вывод, что многие продукты экспрессии Эпштейн-Барр вирусного генома обладают трансформирующими свойствами, а именно EBNA1, EBNA2, EBNA3, LMP1, LMP2A и LMP2B белки, а также EBERs (Эпштейн-Барр вирус кодируемые РНК) и BARTs — продукты транскрипции специфической области вирусного генома [1, p. 987—1001].
Злокачественными новообразования, ассоциированными с Эпштейн-Барр вирусной инфекцией считаются:
1. Назофарингеальная карцинома
2. Лимфома Беркитта
3. ВИЧ/СПИД-ассоциированная лимфома
4. Посттрансплантационные лимфопролиферативные болезни
5. Назальная T/NK-клеточная лимфома
6. Ходжкинская лимфома
7. Аденокарцинома желудка
8. Лимфоэпителиома-подобный рак
9. Лейомиосаркома
По данным зарубежных авторов, главными доказательствами ассоциации латентной Эпштейн-Барр вирусной инфекции и развитием неопластического процесса являются следующие факты, установленные в ходе исследования пациентов, страдающих вышеуказанными опухолями: моноклональность ДНК ВЭБ в опухолевых клетках, высокий уровень свободной вирусной ДНК в плазме крови пациентов, наличие корреляции между титром антивирусных антител, уровнем свободной вирусной ДНК в плазме пациента и тяжестью заболевания, также повышенный титр антивирусных антител часто предшествует появлению новообразования и остается высоким на момент постановки диагноза. Многие из трасформирующих эффектов Эпштейн-Барр вирусных белков и РНК продемонстрированы на животных моделях и культурах клеток [1, p. 987—1001].
Заключение: в настоящее время проблема вирус-ассоциированного онкогенеза очень актуальна и представляет научный интерес для многих специалистов-вирусологов, эпидемиологов и онкологов. В связи с высокой распространенностью Эпштейн-Барр вирусной инфекции среди мирового населения и ее возможными последствиями с исходом в онкологические заболевания особенно важным является вопрос о разработке специфической профилактики. Решением данной проблемы в настоящее время занимаются многие научно-исследовательские центры по всему миру при активном участии ВОЗ. Успешная разработка эффективной и безопасной вакцины против Эпштейн-Барр вируса, как ожидается, приведет не только к снижению заболеваемости острыми формами этой инфекции, но и к снижению заболеваемости Эпштейн-Барр вирус-ассоциированными опухолями по всему миру.
Список литературы:
1.Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis. Edited by Ann Arvin, Gabriella Campadelli-Fiume, Edward Mocarski, Patrick S. Moore, Bernard Roizman, Richard Whitley, and Koichi Yamanishi. Cambridge: Cambridge University Press, 2007:1432
2.Hiroshi K Makoto M, Yumi Y, Kiyotaka K, Koji K, Seiji K, Takaharu M, Tsuneo M. Quantitative Analysis of Epstein-Barr Virus Load by Using a Real-Time PCR Assay. J Clin Microbiol. 1999 January; 37(1): 132—136.
отправлен участнику
Комментарии (1)
Оставить комментарий